

Development of the mouse dermal adipose layer occurs independently of subcutaneous adipose tissue and is marked by restricted early expression of FABP4
- PMID: 23555789
- PMCID: PMC3608551
- DOI: 10.1371/journal.pone.0059811
Development of the mouse dermal adipose layer occurs independently of subcutaneous adipose tissue and is marked by restricted early expression of FABP4
Abstract
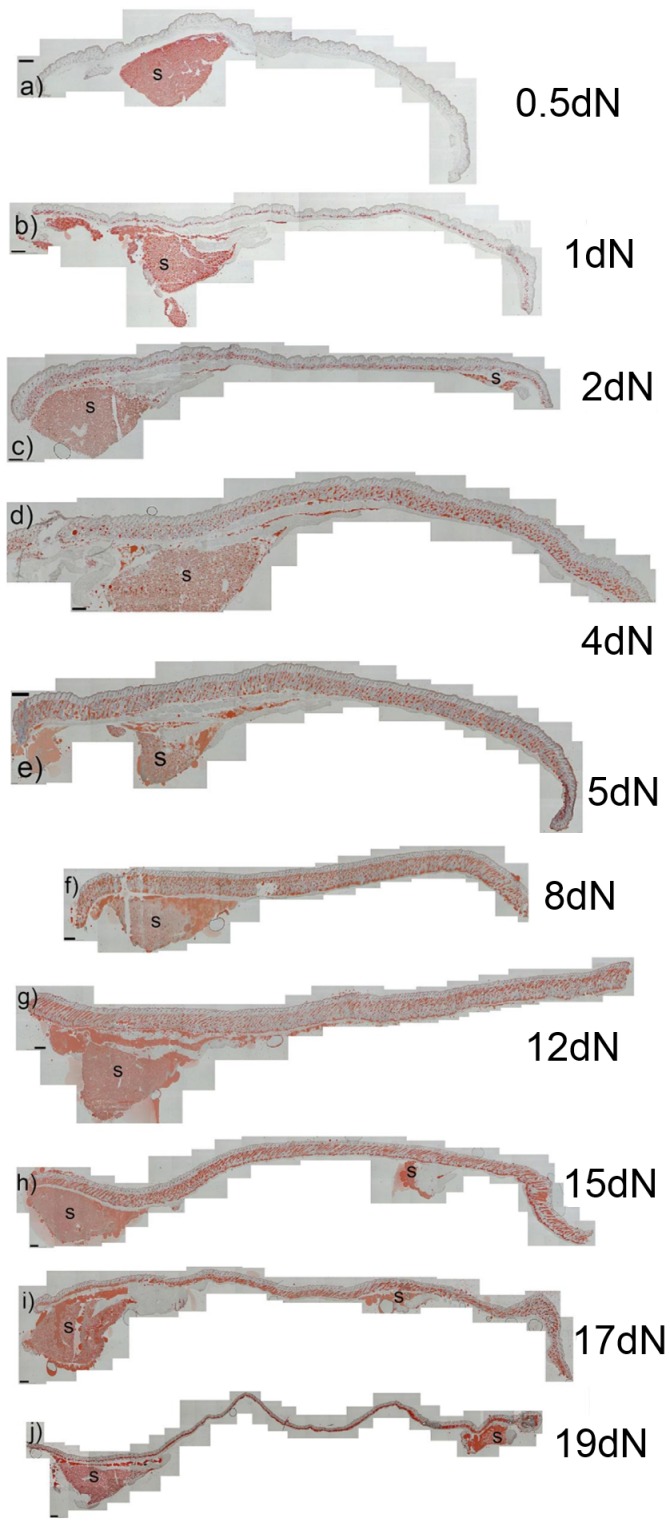
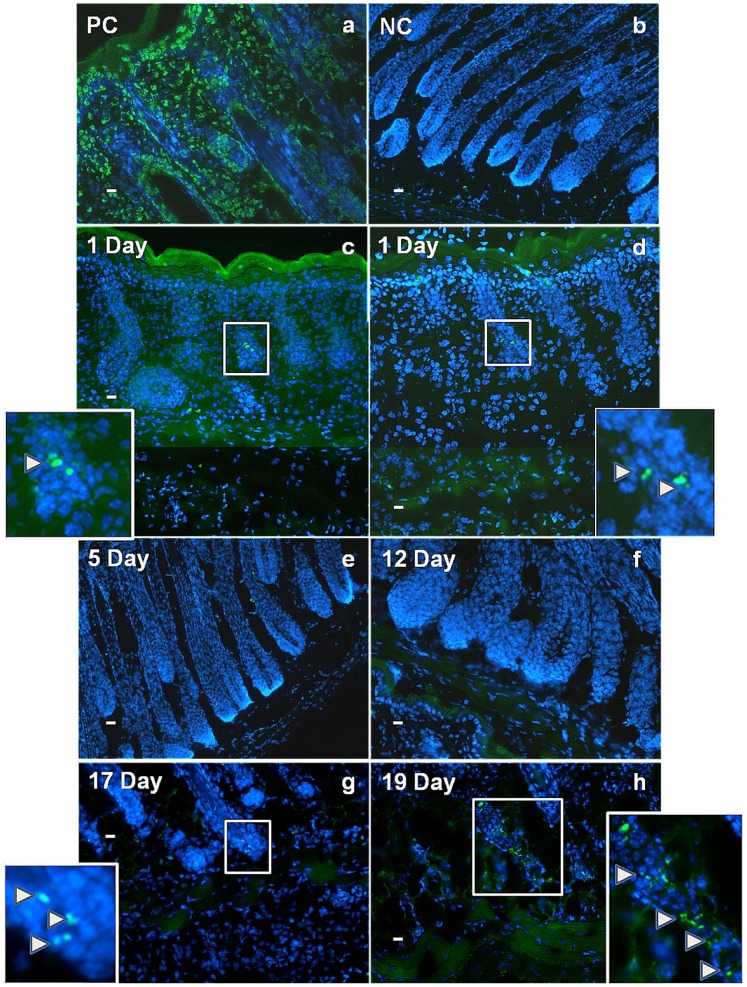
The laboratory mouse is a key animal model for studies of adipose biology, metabolism and disease, yet the developmental changes that occur in tissues and cells that become the adipose layer in mouse skin have received little attention. Moreover, the terminology around this adipose body is often confusing, as frequently no distinction is made between adipose tissue within the skin, and so called subcutaneous fat. Here adipocyte development in mouse dorsal skin was investigated from before birth to the end of the first hair follicle growth cycle. Using Oil Red O staining, immunohistochemistry, quantitative RT-PCR and TUNEL staining we confirmed previous observations of a close spatio-temporal link between hair follicle development and the process of adipogenesis. However, unlike previous studies, we observed that the skin adipose layer was created from cells within the lower dermis. By day 16 of embryonic development (e16) the lower dermis was demarcated from the upper dermal layer, and commitment to adipogenesis in the lower dermis was signalled by expression of FABP4, a marker of adipocyte differentiation. In mature mice the skin adipose layer is separated from underlying subcutaneous adipose tissue by the panniculus carnosus. We observed that the skin adipose tissue did not combine or intermix with subcutaneous adipose tissue at any developmental time point. By transplanting skin isolated from e14.5 mice (prior to the start of adipogenesis), under the kidney capsule of adult mice, we showed that skin adipose tissue develops independently and without influence from subcutaneous depots. This study has reinforced the developmental link between hair follicles and skin adipocyte biology. We argue that because skin adipocytes develop from cells within the dermis and independently from subcutaneous adipose tissue, that it is accurately termed dermal adipose tissue and that, in laboratory mice at least, it represents a separate adipose depot.
Conflict of interest statement
Figures






Similar articles
-
Regulated in Development and DNA Damage Responses 1 Prevents Dermal Adipocyte Differentiation and Is Required for Hair Cycle-Dependent Dermal Adipose Expansion.J Invest Dermatol. 2020 Sep;140(9):1698-1705.e1. doi: 10.1016/j.jid.2019.12.033. Epub 2020 Feb 4. J Invest Dermatol. 2020. PMID: 32032578 Free PMC article.
-
C/EBPalpha identifies differentiating preadipocytes around hair follicles in foetal and neonatal rat and mouse skin.Exp Dermatol. 2008 Aug;17(8):675-80. doi: 10.1111/j.1600-0625.2007.00689.x. Epub 2008 Mar 4. Exp Dermatol. 2008. PMID: 18328085
-
Defining dermal adipose tissue.Exp Dermatol. 2014 Sep;23(9):629-31. doi: 10.1111/exd.12450. Exp Dermatol. 2014. PMID: 24841073 Free PMC article. Review.
-
Dermal white adipose tissue: Development and impact on hair follicles, skin defense, and fibrosis.FASEB J. 2024 Sep 30;38(18):e70047. doi: 10.1096/fj.202400653R. FASEB J. 2024. PMID: 39292527 Review.
-
Hair follicles' transit-amplifying cells govern concurrent dermal adipocyte production through Sonic Hedgehog.Genes Dev. 2016 Oct 15;30(20):2325-2338. doi: 10.1101/gad.285429.116. Epub 2016 Nov 2. Genes Dev. 2016. PMID: 27807033 Free PMC article.
Cited by
-
Anatomy, Histology, and Embryonic Origin of Adipose Tissue: Insights to Understand Adipose Tissue Homofunctionality in Regeneration and Therapies.Adv Exp Med Biol. 2025;1474:53-78. doi: 10.1007/5584_2024_801. Adv Exp Med Biol. 2025. PMID: 39107527 Review.
-
The Modulatable Stem Cell Niche: Tissue Interactions during Hair and Feather Follicle Regeneration.J Mol Biol. 2016 Apr 10;428(7):1423-40. doi: 10.1016/j.jmb.2015.07.009. Epub 2015 Jul 18. J Mol Biol. 2016. PMID: 26196442 Free PMC article. Review.
-
Skin aging: are adipocytes the next target?Aging (Albany NY). 2016 Jul;8(7):1457-69. doi: 10.18632/aging.100999. Aging (Albany NY). 2016. PMID: 27434510 Free PMC article. Review.
-
Functional histology of the skin in the subterranean African giant mole-rat: thermal windows are determined solely by pelage characteristics.PeerJ. 2020 Apr 8;8:e8883. doi: 10.7717/peerj.8883. eCollection 2020. PeerJ. 2020. PMID: 32296606 Free PMC article.
-
Effect of All-trans Retinoic Acid on Panniculus Carnosus Muscle Regeneration in Fetal Mouse Wound Healing.Plast Reconstr Surg Glob Open. 2022 Sep 28;10(9):e4533. doi: 10.1097/GOX.0000000000004533. eCollection 2022 Sep. Plast Reconstr Surg Glob Open. 2022. PMID: 36187276 Free PMC article.
References
-
- Cinti S (2007) Nutrition and Health. In: Fantuzzi G, Mazzone T, editors. Adipose Tissue and Adipokines in Health and Disease. Humana Press Incorporated, Totowa, New Jersey. 3–19.
-
- Gesta S, Tseng YH, Kahn CR (2007) Developmental origin of fat: tracking obesity to its source. Cell 131: 242–256. - PubMed
-
- Billon N, Monteiro MC, Dani C (2008) Developmental origin of adipocytes: new insights into a pending question. Biology of the cell 100: 563–575. - PubMed
-
- Billon N, Dani C (2012) Developmental origins of the adipocyte lineage: new insights from genetics and genomics studies. Stem cell reviews 8: 55–66. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
