

BANK1 and BLK act through phospholipase C gamma 2 in B-cell signaling
- PMID: 23555801
- PMCID: PMC3608554
- DOI: 10.1371/journal.pone.0059842
BANK1 and BLK act through phospholipase C gamma 2 in B-cell signaling
Abstract
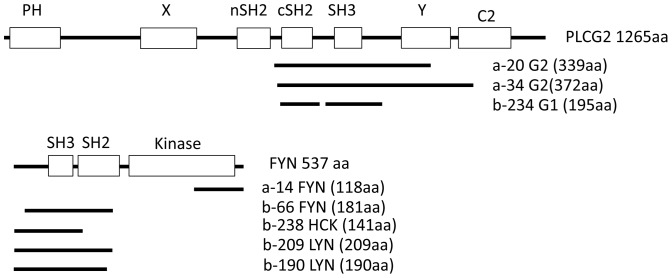
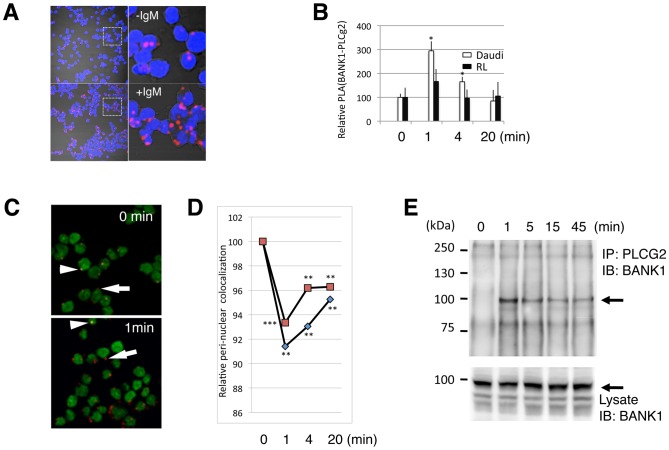
The B cell adaptor protein with ankyrin repeats (BANK1) and the B lymphoid tyrosine kinase (BLK) have been genetically associated with autoimmunity. The proteins of these genes interact physically and work in concert during B-cell signaling. Little is know about their interactions with other B-cell signaling molecules or their role in the process. Using yeast two hybrid (Y2H) we sought for factors that interact with BANK1. We found that the molecular switch PLCg2 interacts with BANK1 and that the interaction is promoted by B-cell receptor (BCR) stimulation. We found further that the kinase activity of BLK enhanced BANK1- PLCg2 binding and that the interaction was suppressed upon BLK depletion. Immunoprecipitation and mutational analysis demonstrated that the interaction between BANK1 and PLCg2 was dependent on specific tyrosine and proline residues on the adaptor protein. Our results provide new information important to understand the role of these two genes in basic B-cell physiology and immune-related diseases.
Conflict of interest statement
Figures




References
-
- Chan VW, Lowell CA, DeFranco AL (1998) Defective negative regulation of antigen receptor signaling in Lyn-deficient B lymphocytes. Curr Biol 8: 545–553. - PubMed
-
- Sato S, Hasegawa M, Fujimoto M, Tedder TF, Takehara K (2000) Quantitative genetic variation in CD19 expression correlates with autoimmunity. J Immunol 165: 6635–6643. - PubMed
-
- Majeti R, Xu Z, Parslow TG, Olson JL, Daikh DI, et al. (2000) An inactivating point mutation in the inhibitory wedge of CD45 causes lymphoproliferation and autoimmunity. Cell 103: 1059–1070. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
