

PAPP5 is involved in the tetrapyrrole mediated plastid signalling during chloroplast development
- PMID: 23555952
- PMCID: PMC3612061
- DOI: 10.1371/journal.pone.0060305
PAPP5 is involved in the tetrapyrrole mediated plastid signalling during chloroplast development
Abstract
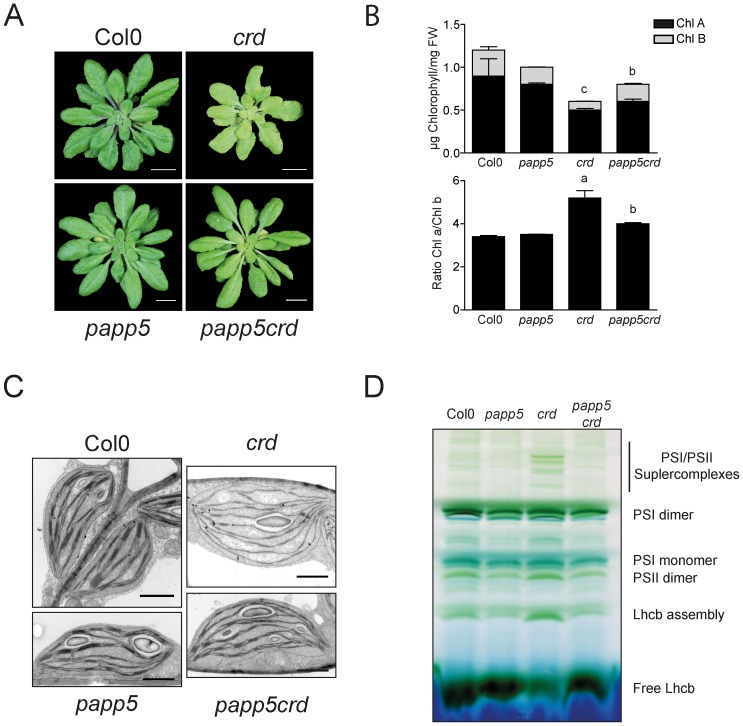
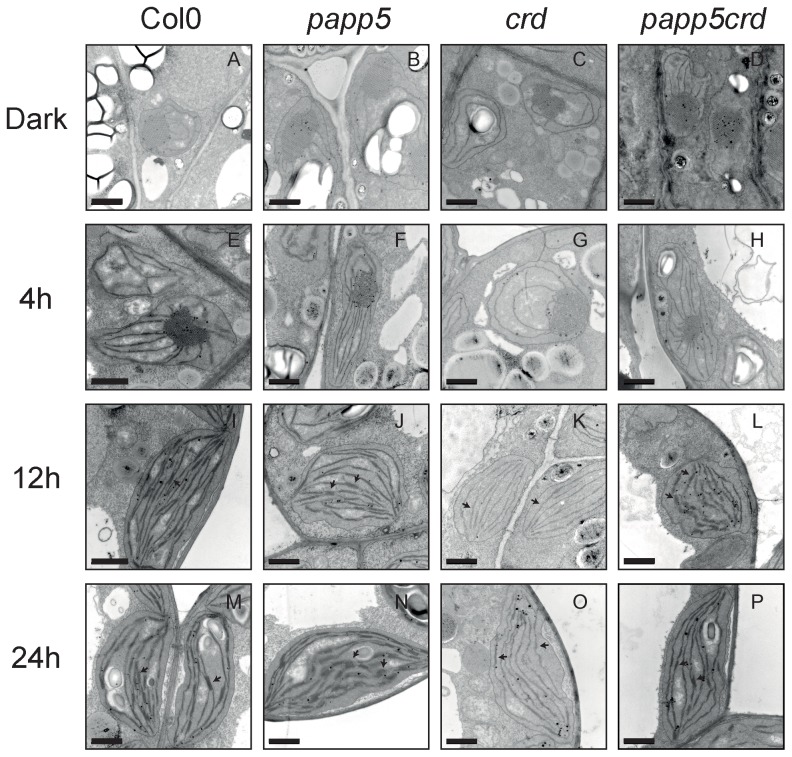
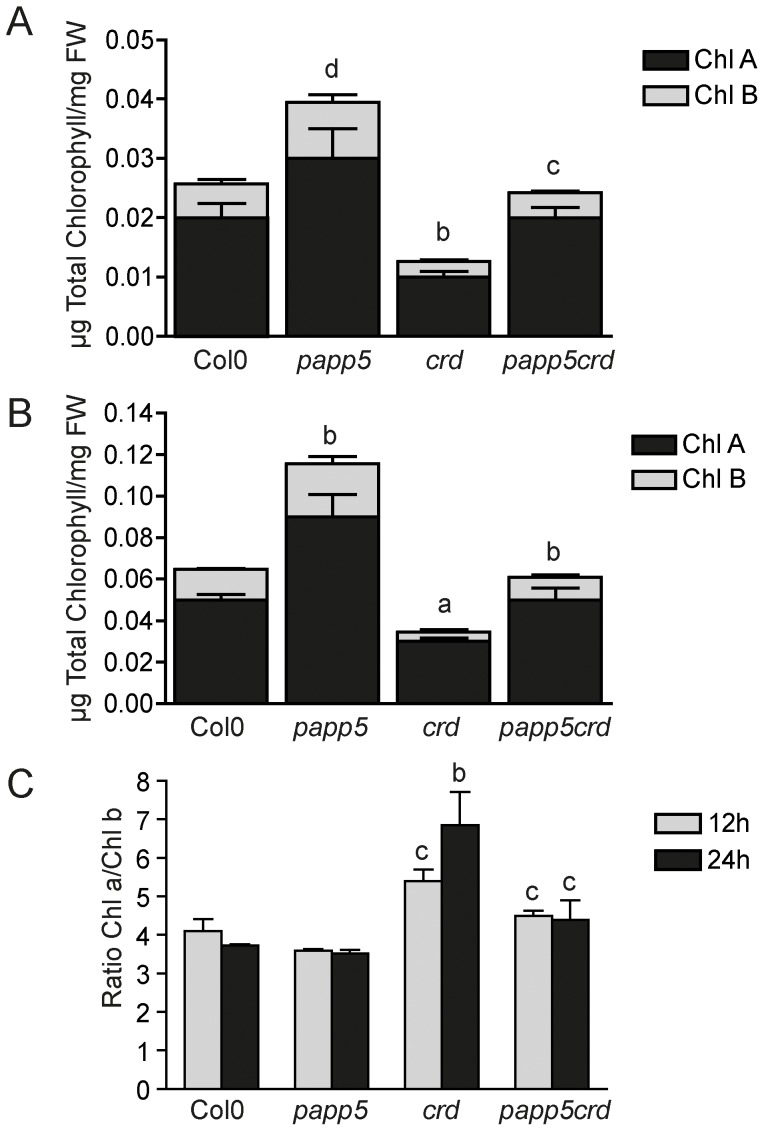
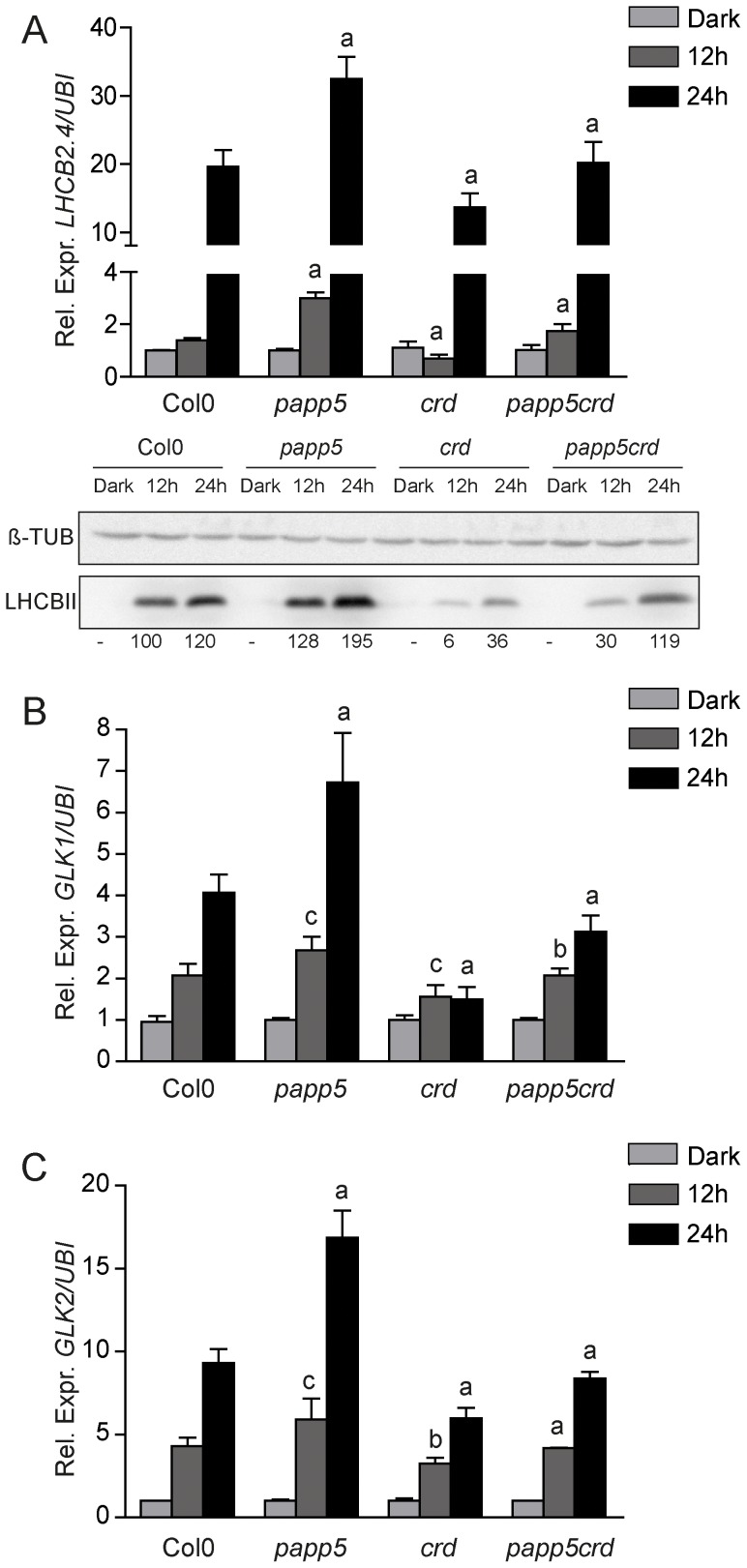
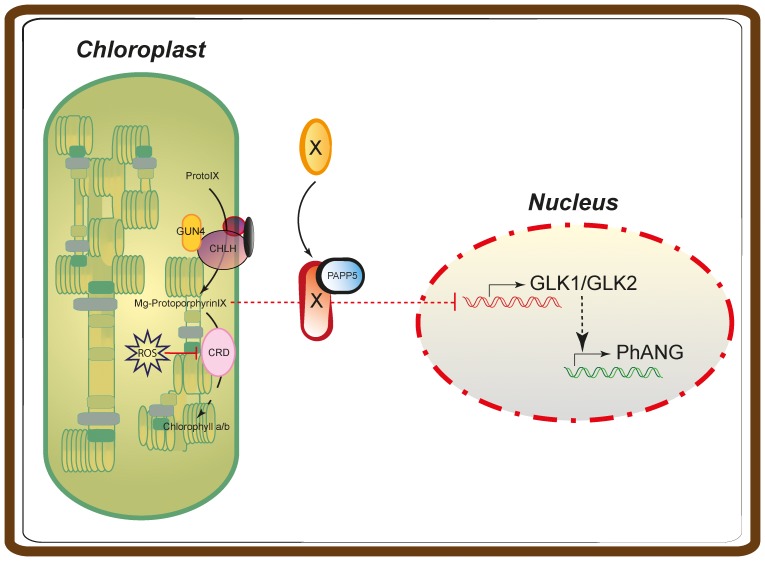
The initiation of chloroplast development in the light is dependent on nuclear encoded components. The nuclear genes encoding key components in the photosynthetic machinery are regulated by signals originating in the plastids. These plastid signals play an essential role in the regulation of photosynthesis associated nuclear genes (PhANGs) when proplastids develop into chloroplasts. One of the plastid signals is linked to the tetrapyrrole biosynthesis and accumulation of the intermediates the Mg-ProtoIX and its methyl ester Mg-ProtoIX-ME. Phytochrome-Associated Protein Phosphatase 5 (PAPP5) was isolated in a previous study as a putative Mg-ProtoIX interacting protein. In order to elucidate if there is a biological link between PAPP5 and the tetrapyrrole mediated signal we generated double mutants between the Arabidopsis papp5 and the crd mutants. The crd mutant over-accumulates Mg-ProtoIX and Mg-ProtoIX-ME and the tetrapyrrole accumulation triggers retrograde signalling. The crd mutant exhibits repression of PhANG expression, altered chloroplast morphology and a pale phenotype. However, in the papp5crd double mutant, the crd phenotype is restored and papp5crd accumulated wild type levels of chlorophyll, developed proper chloroplasts and showed normal induction of PhANG expression in response to light. Tetrapyrrole feeding experiments showed that PAPP5 is required to respond correctly to accumulation of tetrapyrroles in the cell and that PAPP5 is most likely a component in the plastid signalling pathway down stream of the tetrapyrrole Mg-ProtoIX/Mg-ProtoIX-ME. Inhibition of phosphatase activity phenocopied the papp5crd phenotype in the crd single mutant demonstrating that PAPP5 phosphatase activity is essential to mediate the retrograde signal and to suppress PhANG expression in the crd mutant. Thus, our results suggest that PAPP5 receives an inbalance in the tetrapyrrole biosynthesis through the accumulation of Mg-ProtoIX and acts as a negative regulator of PhANG expression during chloroplast biogenesis and development.
Conflict of interest statement
Figures




References
-
- Chen M, Chory J, Fankhauser C (2004) Light signal transduction in higher plants. Annu Rev Genet 38: 87–117. - PubMed
-
- Leon P, Arroyo A, Mackenzie S (1998) Nuclear Control of Plastid and Mitochondrial Development in Higher Plants. Annu Rev Plant Physiol Plant Mol Biol 49: 453–480. - PubMed
-
- Oelmuller R, Levitan I, Bergfeld R, Rajasekhar V, Mohr H (1986) Expression of nuclear genes is affected by treatments acting on the plastids. Planta 168: 482–492. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
