

Arabidopsis DEAD-box RNA helicase UAP56 interacts with both RNA and DNA as well as with mRNA export factors
- PMID: 23555998
- PMCID: PMC3608606
- DOI: 10.1371/journal.pone.0060644
Arabidopsis DEAD-box RNA helicase UAP56 interacts with both RNA and DNA as well as with mRNA export factors
Abstract
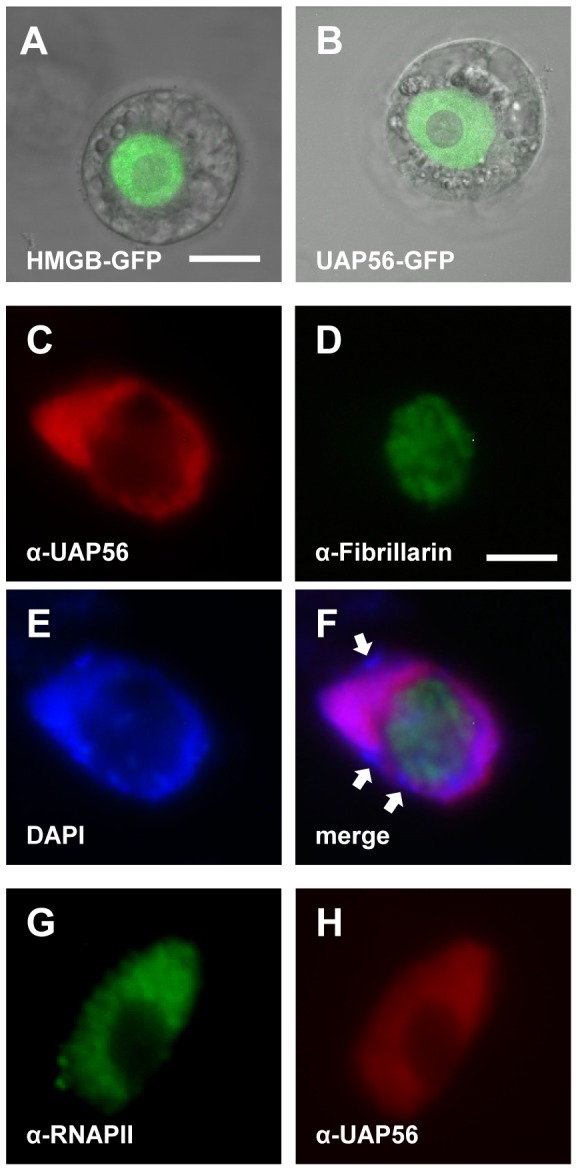
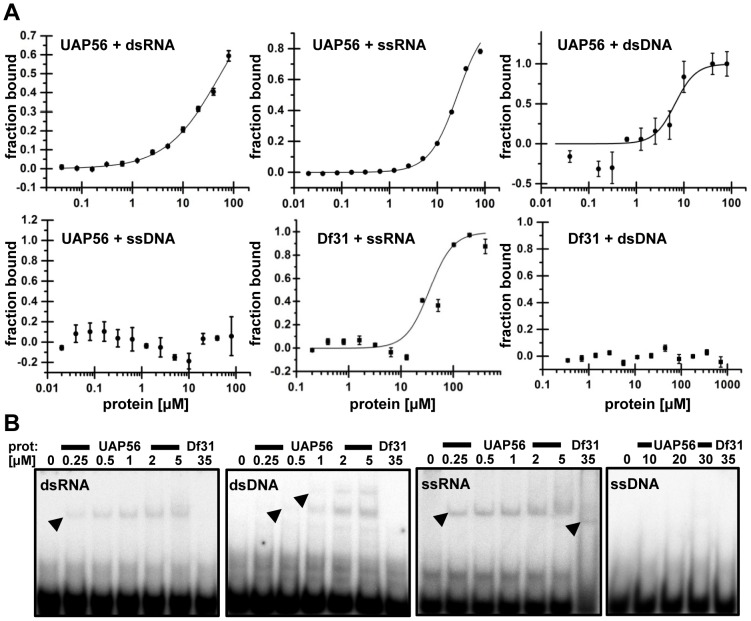
The DEAD-box protein UAP56 (U2AF65-associcated protein) is an RNA helicase that in yeast and metazoa is critically involved in mRNA splicing and export. In Arabidopsis, two adjacent genes code for an identical UAP56 protein, and both genes are expressed. In case one of the genes is inactivated by a T-DNA insertion, wild type transcript level is maintained by the other intact gene. In contrast to other organisms that are severely affected by elevated UAP56 levels, Arabidopsis plants that overexpress UAP56 have wild type appearance. UAP56 localises predominantly to euchromatic regions of Arabidopsis nuclei, and associates with genes transcribed by RNA polymerase II independently from the presence of introns, while it is not detected at non-transcribed loci. Biochemical characterisation revealed that in addition to ssRNA and dsRNA, UAP56 interacts with dsDNA, but not with ssDNA. Moreover, the enzyme displays ATPase activity that is stimulated by RNA and dsDNA and it has ATP-dependent RNA helicase activity unwinding dsRNA, whereas it does not unwind dsDNA. Protein interaction studies showed that UAP56 directly interacts with the mRNA export factors ALY2 and MOS11, suggesting that it is involved in mRNA export from plant cell nuclei.
Conflict of interest statement
Figures





References
-
- Pyle AM (2008) Unwinding mechanisms of RNA and DNA helicases. Ann Rev Biophys 37: 317–336. - PubMed
-
- Linder P, Jankowsky E (2011) From unwinding to clamping - the DEAD box RNA helicase family. Nat Rev Mol Cell Biol 12: 505–516. - PubMed
-
- Fleckner J, Zhang M, Valcarcel J, Green MR (1997) U2AF65 recruits a novel human DEAD box protein required for the U2 snRNP-branchpoint interaction. Genes Dev 11: 1864–1872. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
