

Deletion of the last five C-terminal amino acid residues of connexin43 leads to lethal ventricular arrhythmias in mice without affecting coupling via gap junction channels
- PMID: 23558439
- PMCID: PMC3678986
- DOI: 10.1007/s00395-013-0348-y
Deletion of the last five C-terminal amino acid residues of connexin43 leads to lethal ventricular arrhythmias in mice without affecting coupling via gap junction channels
Abstract
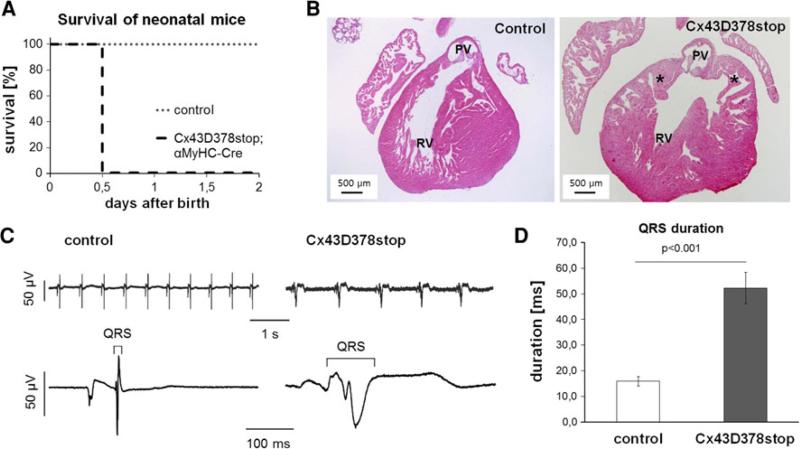
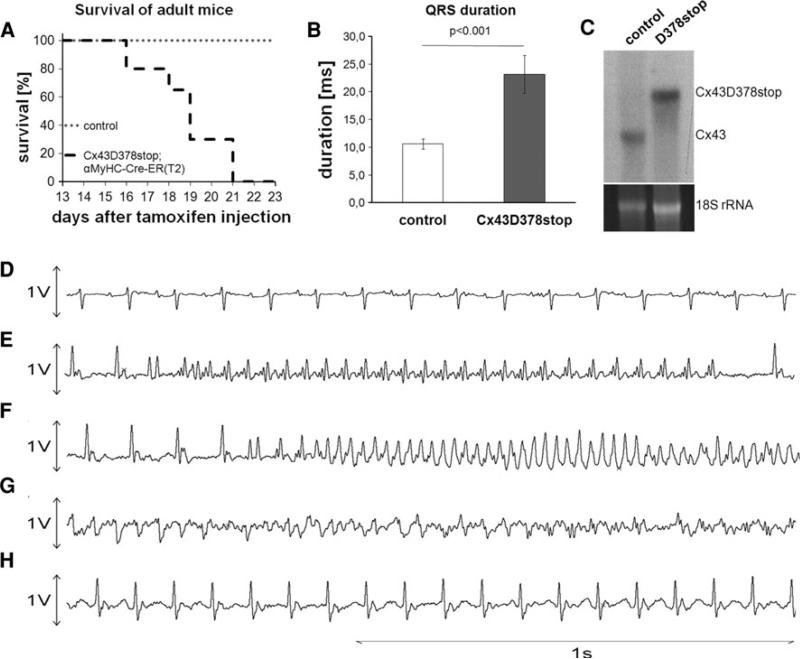
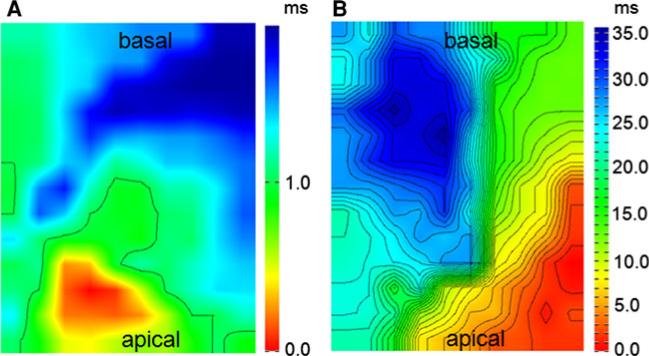
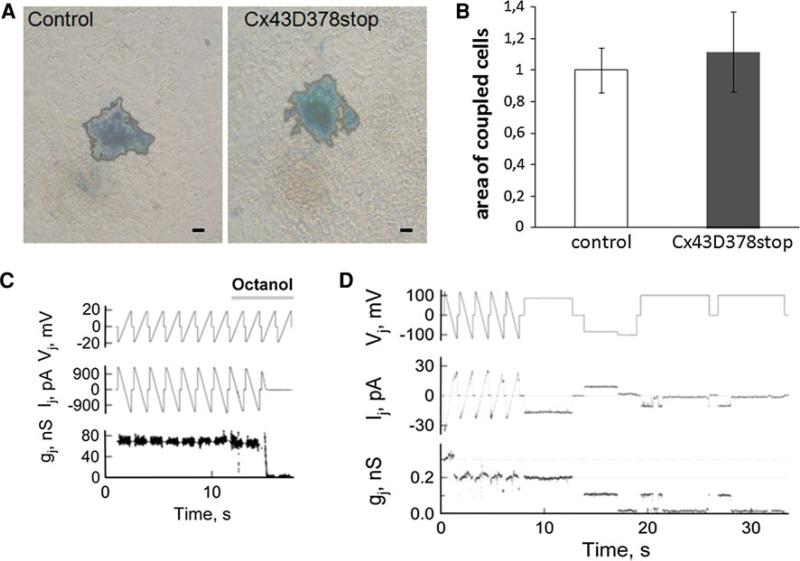
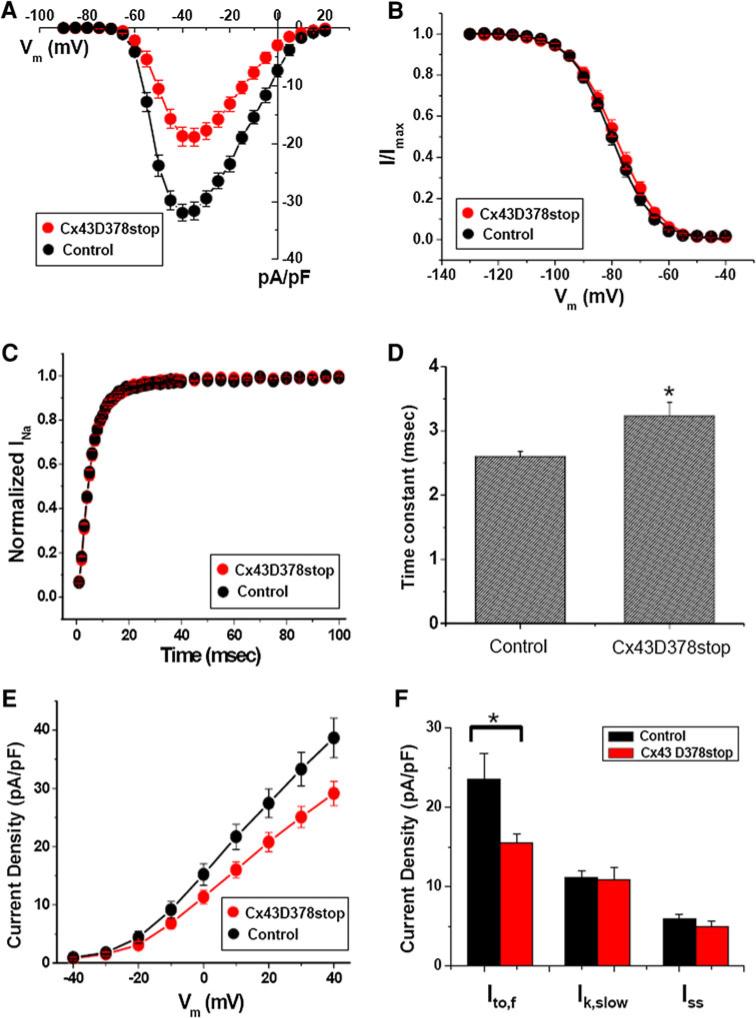
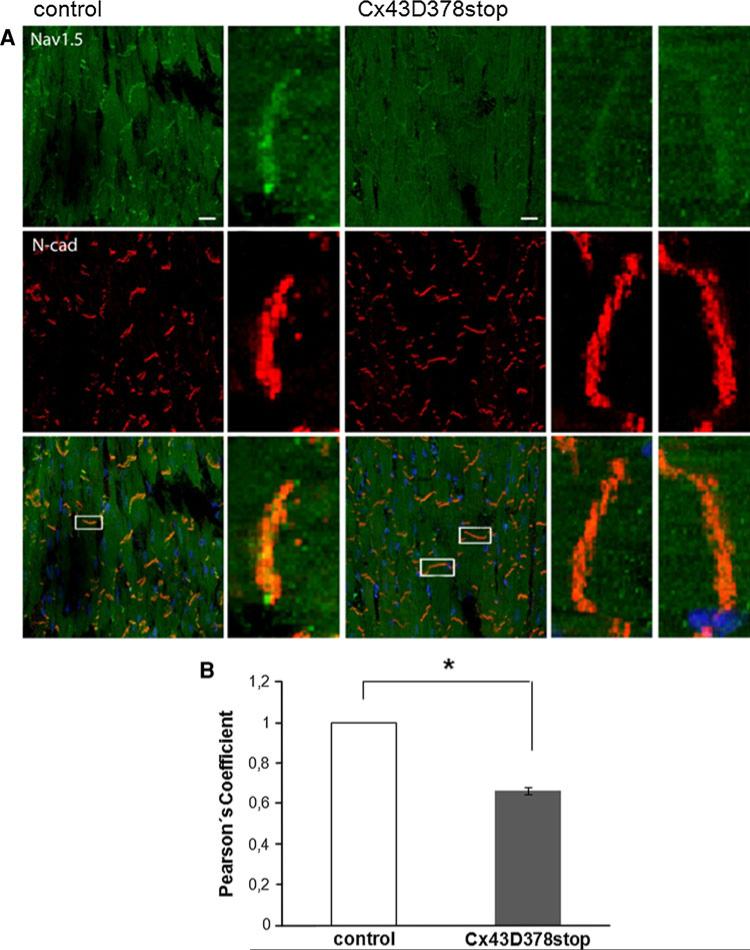
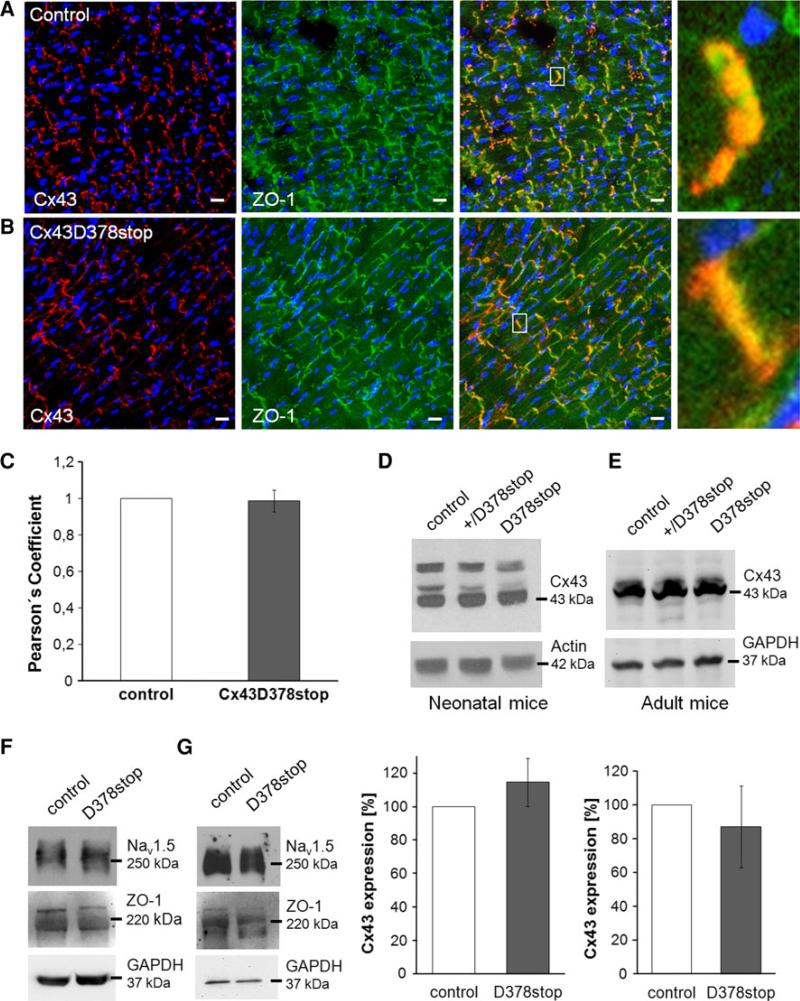
The cardiac intercalated disc harbors mechanical and electrical junctions as well as ion channel complexes mediating propagation of electrical impulses. Cardiac connexin43 (Cx43) co-localizes and interacts with several of the proteins located at intercalated discs in the ventricular myocardium. We have generated conditional Cx43D378stop mice lacking the last five C-terminal amino acid residues, representing a binding motif for zonula occludens protein-1 (ZO-1), and investigated the functional consequences of this mutation on cardiac physiology and morphology. Newborn and adult homozygous Cx43D378stop mice displayed markedly impaired and heterogeneous cardiac electrical activation properties and died from severe ventricular arrhythmias. Cx43 and ZO-1 were co-localized at intercalated discs in Cx43D378stop hearts, and the Cx43D378stop gap junction channels showed normal coupling properties. Patch clamp analyses of isolated adult Cx43D378stop cardiomyocytes revealed a significant decrease in sodium and potassium current densities. Furthermore, we also observed a significant loss of Nav1.5 protein from intercalated discs in Cx43D378stop hearts. The phenotypic lethality of the Cx43D378stop mutation was very similar to the one previously reported for adult Cx43 deficient (Cx43KO) mice. Yet, in contrast to Cx43KO mice, the Cx43 gap junction channel was still functional in the Cx43D378stop mutant. We conclude that the lethality of Cx43D378stop mice is independent of the loss of gap junctional intercellular communication, but most likely results from impaired cardiac sodium and potassium currents. The Cx43D378stop mice reveal for the first time that Cx43 dependent arrhythmias can develop by mechanisms other than impairment of gap junction channel function.
Figures
Similar articles
-
Super-resolution imaging reveals that loss of the C-terminus of connexin43 limits microtubule plus-end capture and NaV1.5 localization at the intercalated disc.Cardiovasc Res. 2014 Nov 1;104(2):371-81. doi: 10.1093/cvr/cvu195. Epub 2014 Aug 18. Cardiovasc Res. 2014. PMID: 25139742 Free PMC article.
-
Potassium channels in the Cx43 gap junction perinexus modulate ephaptic coupling: an experimental and modeling study.Pflugers Arch. 2016 Oct;468(10):1651-61. doi: 10.1007/s00424-016-1861-2. Epub 2016 Aug 11. Pflugers Arch. 2016. PMID: 27510622 Free PMC article.
-
Silencing of desmoplakin decreases connexin43/Nav1.5 expression and sodium current in HL‑1 cardiomyocytes.Mol Med Rep. 2013 Sep;8(3):780-6. doi: 10.3892/mmr.2013.1594. Epub 2013 Jul 18. Mol Med Rep. 2013. PMID: 23877755
-
Gap junctions - guards of excitability.Biochem Soc Trans. 2015 Jun;43(3):508-12. doi: 10.1042/BST20150059. Biochem Soc Trans. 2015. PMID: 26009199 Review.
-
Intracellular trafficking pathways of Cx43 gap junction channels.Biochim Biophys Acta Biomembr. 2018 Jan;1860(1):40-47. doi: 10.1016/j.bbamem.2017.05.018. Epub 2017 May 30. Biochim Biophys Acta Biomembr. 2018. PMID: 28576298 Free PMC article. Review.
Cited by
-
Pathogenesis and management of Brugada syndrome.Nat Rev Cardiol. 2016 Dec;13(12):744-756. doi: 10.1038/nrcardio.2016.143. Epub 2016 Sep 15. Nat Rev Cardiol. 2016. PMID: 27629507 Review.
-
Mechanisms of Arrhythmias in the Brugada Syndrome.Int J Mol Sci. 2020 Sep 25;21(19):7051. doi: 10.3390/ijms21197051. Int J Mol Sci. 2020. PMID: 32992720 Free PMC article. Review.
-
Arrhythmogenic cardiomyopathy and Brugada syndrome: diseases of the connexome.FEBS Lett. 2014 Apr 17;588(8):1322-30. doi: 10.1016/j.febslet.2014.02.008. Epub 2014 Feb 15. FEBS Lett. 2014. PMID: 24548564 Free PMC article. Review.
-
Connexin 43 dephosphorylation at serine 282 induces spontaneous arrhythmia and increases susceptibility to ischemia/reperfusion injury.Heliyon. 2023 May 1;9(5):e15879. doi: 10.1016/j.heliyon.2023.e15879. eCollection 2023 May. Heliyon. 2023. PMID: 37215881 Free PMC article.
-
Mechanisms of Intracellular Communication in Cancer and Pathogen Spreading.Results Probl Cell Differ. 2024;73:301-326. doi: 10.1007/978-3-031-62036-2_13. Results Probl Cell Differ. 2024. PMID: 39242384 Review.
References
-
- Agah R, Frenkel PA, French BA, Michael LH, Overbeek PA, Schneider MD. Gene recombination in postmitotic cells. Targeted expression of Cre recombinase provokes cardiac-restricted, site-specific rearrangement in adult ventricular muscle in vivo. J Clin Invest. 1997;100:169–179. doi:10.1172/JCI119509. - PMC - PubMed
-
- Asimaki A, Tandri H, Huang H, Halushka MK, Gautam S, Basso C, Thiene G, Tsatsopoulou A, Protonotarios N, McKenna WJ, Calkins H, Saffitz JE. A new diagnostic test for arrhythmogenic right ventricular cardiomyopathy. N Engl J Med. 2009;360:1075–1084. doi:10.1056/NEJMoa0808138. - PubMed
-
- Boengler K, Dodoni G, Rodriguez-Sinovas A, Cabestrero A, Ruiz-Meana M, Gres P, Konietzka I, Lopez-Iglesias C, Garcia-Dorado D, Di Lisa F, Heusch G, Schulz R. Connexin 43 in cardiomyocyte mitochondria and its increase by ischemic preconditioning. Cardiovasc Res. 2005;67:234–244. doi:10.1016/j.cardiores.2005.04.014. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- HL084583/HL/NHLBI NIH HHS/United States
- R01 NS072238/NS/NINDS NIH HHS/United States
- R01NS072238/NS/NINDS NIH HHS/United States
- R01 HL106632/HL/NHLBI NIH HHS/United States
- R01 HL106632-01/HL/NHLBI NIH HHS/United States
- R01 GM57691-13/GM/NIGMS NIH HHS/United States
- R01 HL083422/HL/NHLBI NIH HHS/United States
- HL083422/HL/NHLBI NIH HHS/United States
- R01 HL084464/HL/NHLBI NIH HHS/United States
- R01 HL084583/HL/NHLBI NIH HHS/United States
- R01HL084464/HL/NHLBI NIH HHS/United States
- R01 GM057691/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
