

Drosophila H1 regulates the genetic activity of heterochromatin by recruitment of Su(var)3-9
- PMID: 23559249
- PMCID: PMC3756538
- DOI: 10.1126/science.1234654
Drosophila H1 regulates the genetic activity of heterochromatin by recruitment of Su(var)3-9
Abstract
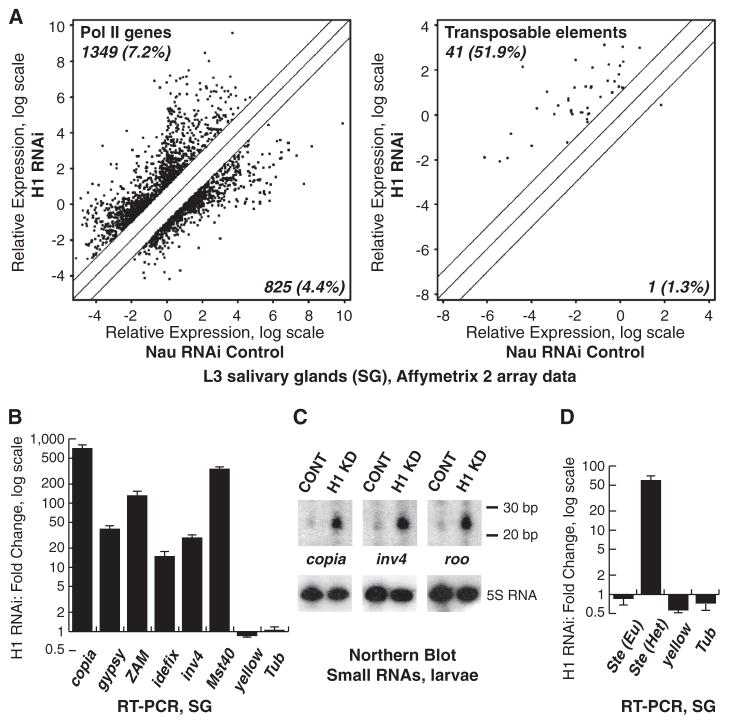
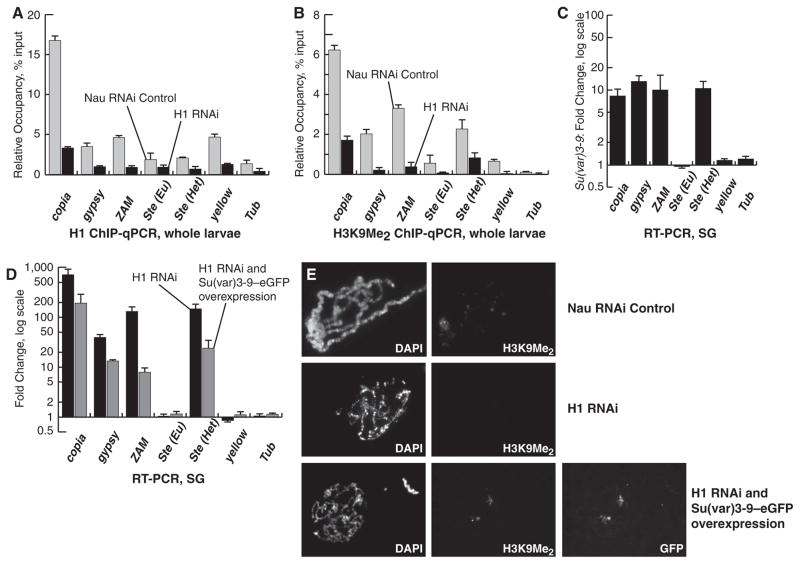
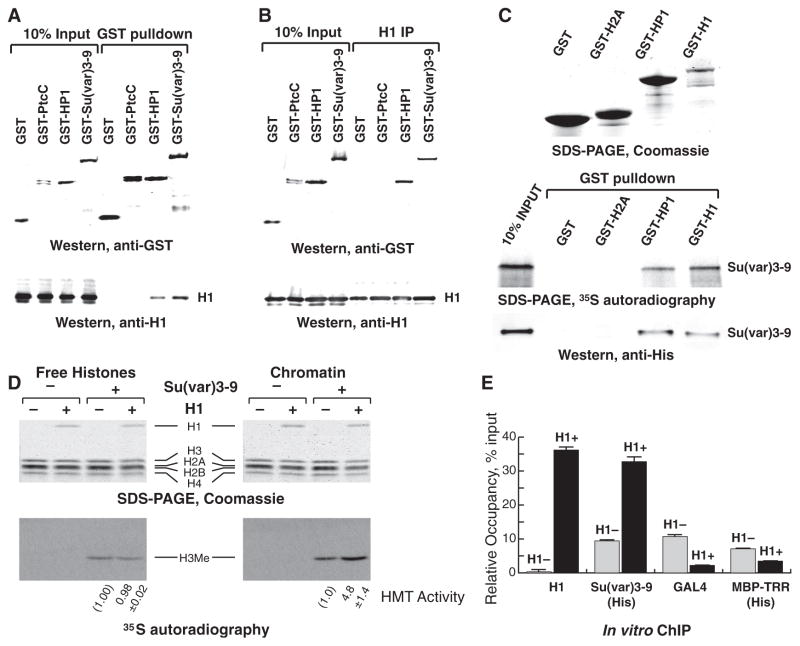
Eukaryotic genomes harbor transposable elements and other repetitive sequences that must be silenced. Small RNA interference pathways play a major role in their repression. Here, we reveal another mechanism for silencing these sequences in Drosophila. Depleting the linker histone H1 in vivo leads to strong activation of these elements. H1-mediated silencing occurs in combination with the heterochromatin-specific histone H3 lysine 9 methyltransferase Su(var)3-9. H1 physically interacts with Su(var)3-9 and recruits it to chromatin in vitro, which promotes H3 methylation. We propose that H1 plays a key role in silencing by tethering Su(var)3-9 to heterochromatin. The tethering function of H1 adds to its established role as a regulator of chromatin compaction and accessibility.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
