

Can insects develop resistance to insect pathogenic fungi?
- PMID: 23560083
- PMCID: PMC3613352
- DOI: 10.1371/journal.pone.0060248
Can insects develop resistance to insect pathogenic fungi?
Erratum in
- PLoS One. 2014;9(1). doi:10.1371/annotation/3c61c1d6-7981-4f3e-a690-1ce7a4d89285
Abstract
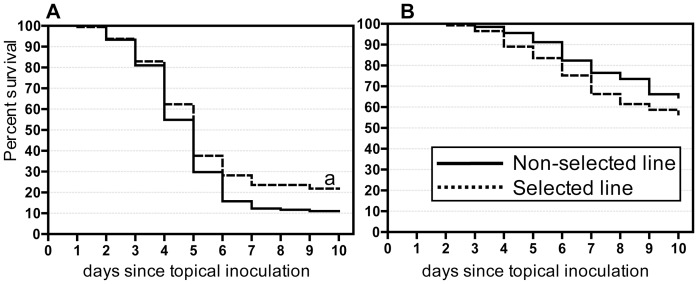
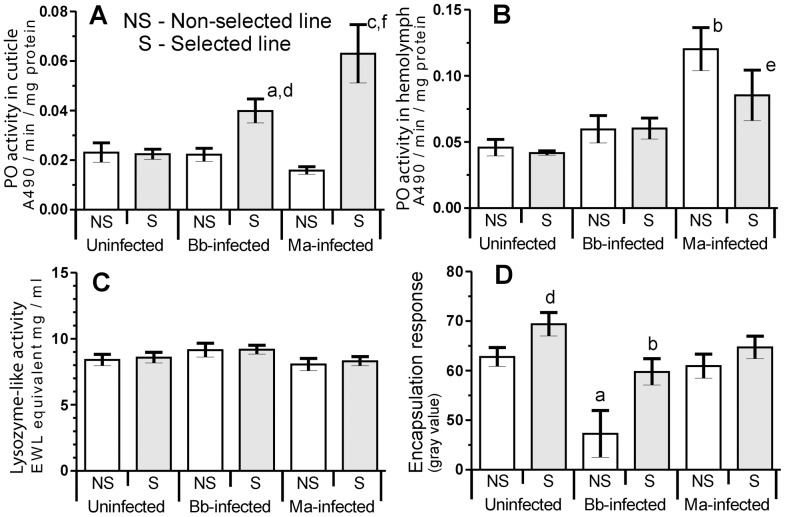
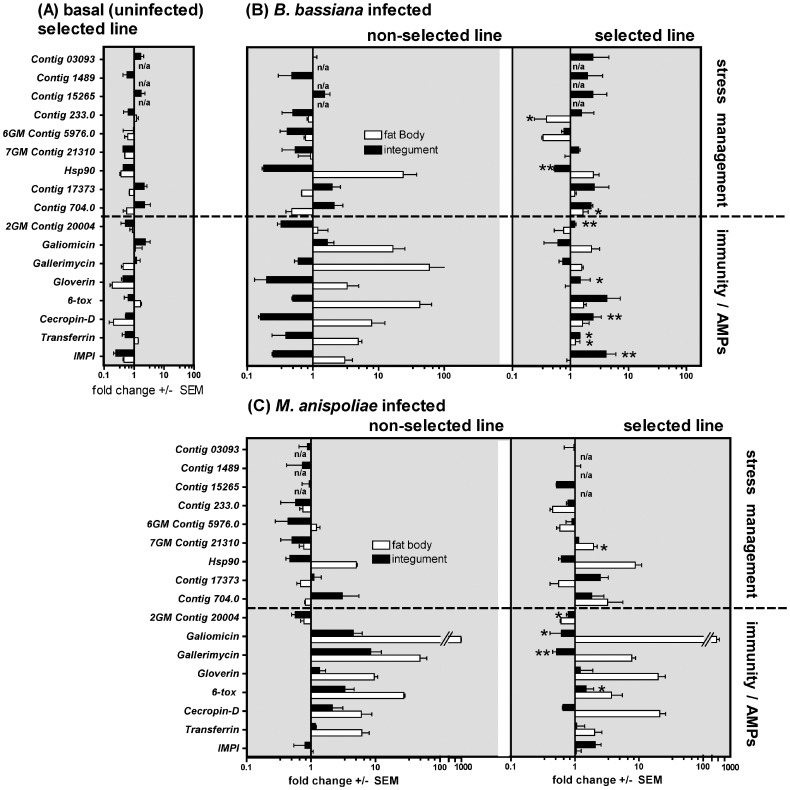
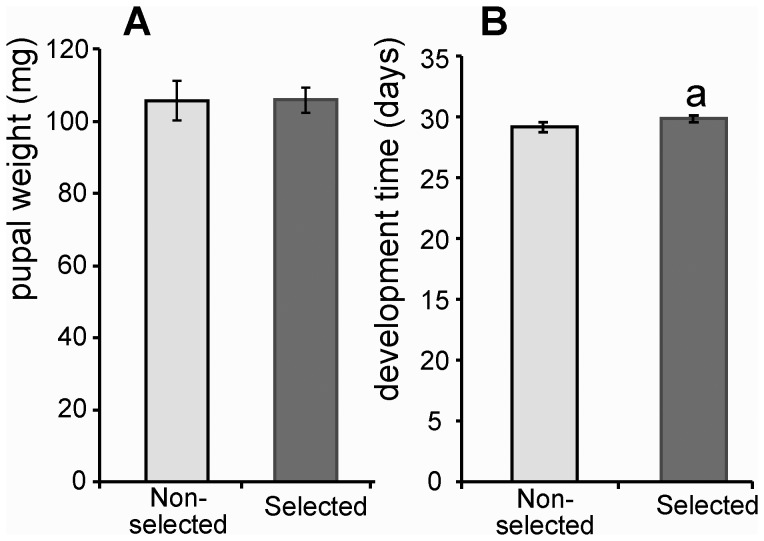
Microevolutionary adaptations and mechanisms of fungal pathogen resistance were explored in a melanic population of the Greater wax moth, Galleria mellonella. Under constant selective pressure from the insect pathogenic fungus Beauveria bassiana, 25(th) generation larvae exhibited significantly enhanced resistance, which was specific to this pathogen and not to another insect pathogenic fungus, Metarhizium anisopliae. Defense and stress management strategies of selected (resistant) and non-selected (susceptible) insect lines were compared to uncover mechanisms underpinning resistance, and the possible cost of those survival strategies. We hypothesize that the insects developed a transgenerationally primed resistance to the fungus B. bassiana, a costly trait that was achieved not by compromising life-history traits but rather by prioritizing and re-allocating pathogen-species-specific augmentations to integumental front-line defenses that are most likely to be encountered by invading fungi. Specifically during B. bassiana infection, systemic immune defenses are suppressed in favour of a more limited but targeted repertoire of enhanced responses in the cuticle and epidermis of the integument (e.g. expression of the fungal enzyme inhibitor IMPI, and cuticular phenoloxidase activity). A range of putative stress-management factors (e.g. antioxidants) is also activated during the specific response of selected insects to B. bassiana but not M. anisopliae. This too occurs primarily in the integument, and probably contributes to antifungal defense and/or helps ameliorate the damage inflicted by the fungus or the host's own immune responses.
Conflict of interest statement
Figures
References
-
- Wraight SP, Butt TM, Galaini-Wraight S, Allee LL, Soper RS, et al. (1990) Germination and infection processes of the entomophthoralean fungus Erynia radicans on the potato leafhopper, Empoasca fabae . Journal of Invertebrate Pathology 56: 157–174.
-
- Hajek AE, St. Leger RJ (1994) Interactions between fungal pathogens and insect hosts. Annual Review of Entomology 39: 293–322.
-
- Yan XD, Scherphof GL, Kamps J (2005) Liposome opsonization. Journal of Liposome Research 15: 109–139. - PubMed
-
- Butt TM, Wraight SP, Galaini-Wraight S, Humber RA, Roberts DW, et al. (1988) Humoral encapsulation of the fungus Erynia radicans (Entomophthorales) by the potato leafhopper, Empoasca fabae (Homoptera: Cicadellidae). Journal of Invertebrate Pathology 52: 49–56.
-
- Cerenius L, Lee BL, Söderhäll K (2008) The proPO-system: pros and cons for its role in invertebrate immunity. Trends in Immunology 29: 263–271. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
