

New components of Drosophila leg development identified through genome wide association studies
- PMID: 23560084
- PMCID: PMC3613359
- DOI: 10.1371/journal.pone.0060261
New components of Drosophila leg development identified through genome wide association studies
Abstract
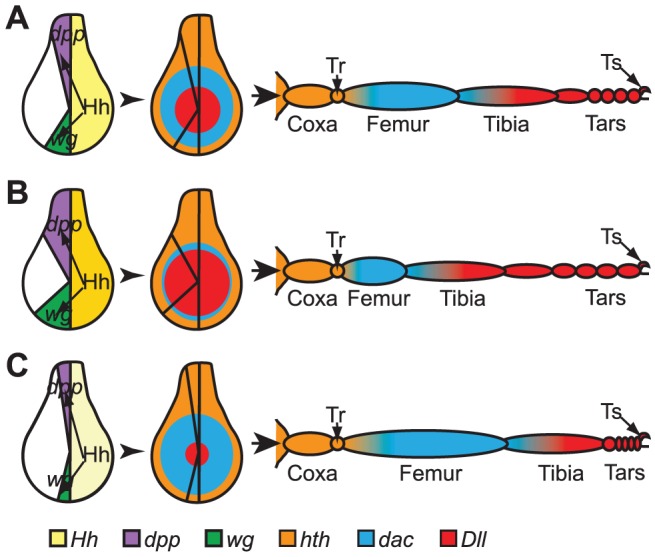
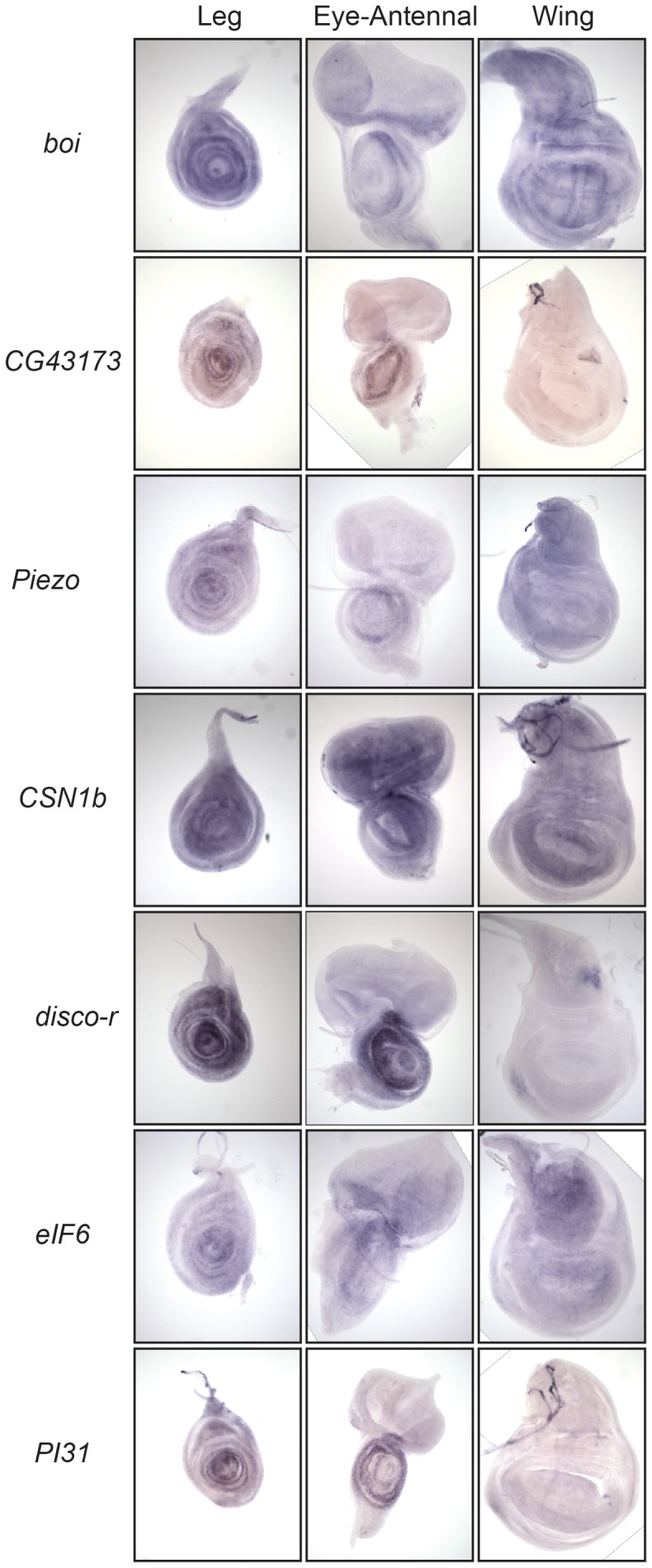
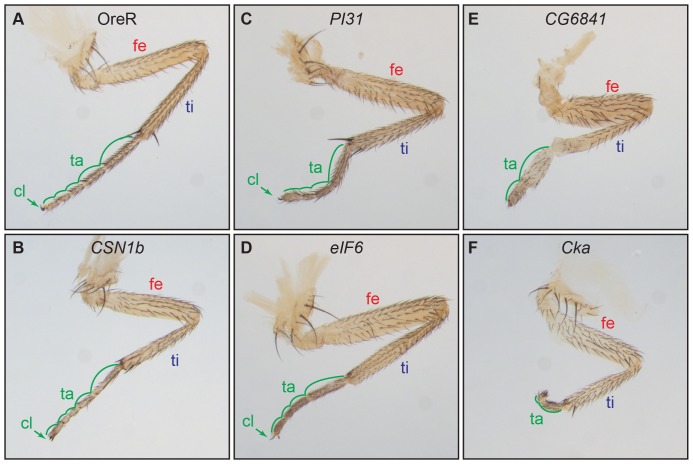
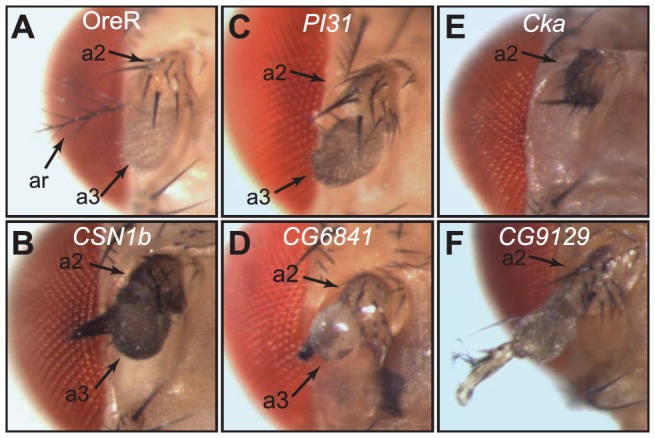
The adult Drosophila melanogaster body develops from imaginal discs, groups of cells set-aside during embryogenesis and expanded in number during larval stages. Specification and development of Drosophila imaginal discs have been studied for many years as models of morphogenesis. These studies are often based on mutations with large developmental effects, mutations that are often lethal in embryos when homozygous. Such forward genetic screens can be limited by factors such as early lethality and genetic redundancy. To identify additional genes and genetic pathways involved in leg imaginal disc development, we employed a Genome Wide Association Study utilizing the natural genetic variation in leg proportionality found in the Drosophila Genetic Reference Panel fly lines. In addition to identifying genes already known to be involved in leg development, we identified several genes involved in pathways that had not previously been linked with leg development. Several of the genes appear to be involved in signaling activities, while others have no known roles at this time. Many of these uncharacterized genes are conserved in mammals, so we can now begin to place these genes into developmental contexts. Interestingly, we identified five genes which, when their function is reduced by RNAi, cause an antenna-to-leg transformation. Our results demonstrate the utility of this approach, integrating the tools of quantitative and molecular genetics to study developmental processes, and provide new insights into the pathways and networks involved in Drosophila leg development.
Conflict of interest statement
Figures
References
-
- Kojima T (2004) The mechanism of Drosophila leg development along the proximodistal axis. Dev Growth Differ 46: 115–129. - PubMed
-
- Morata G (2001) How Drosophila appendages develop. Nat Rev Mol Cell Biol 2: 89–97. - PubMed
-
- Pueyo JI, Couso JP (2005) Parallels between the proximal-distal development of vertebrate and arthropod appendages: homology without an ancestor? Curr Opin Genet Dev 15: 439–446. - PubMed
-
- Panganiban G (2000) Distal-less function during Drosophila appendage and sense organ development. Dev Dyn 218: 554–562. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
