

Apoptosome structure, assembly, and procaspase activation
- PMID: 23561633
- PMCID: PMC3644875
- DOI: 10.1016/j.str.2013.02.024
Apoptosome structure, assembly, and procaspase activation
Abstract
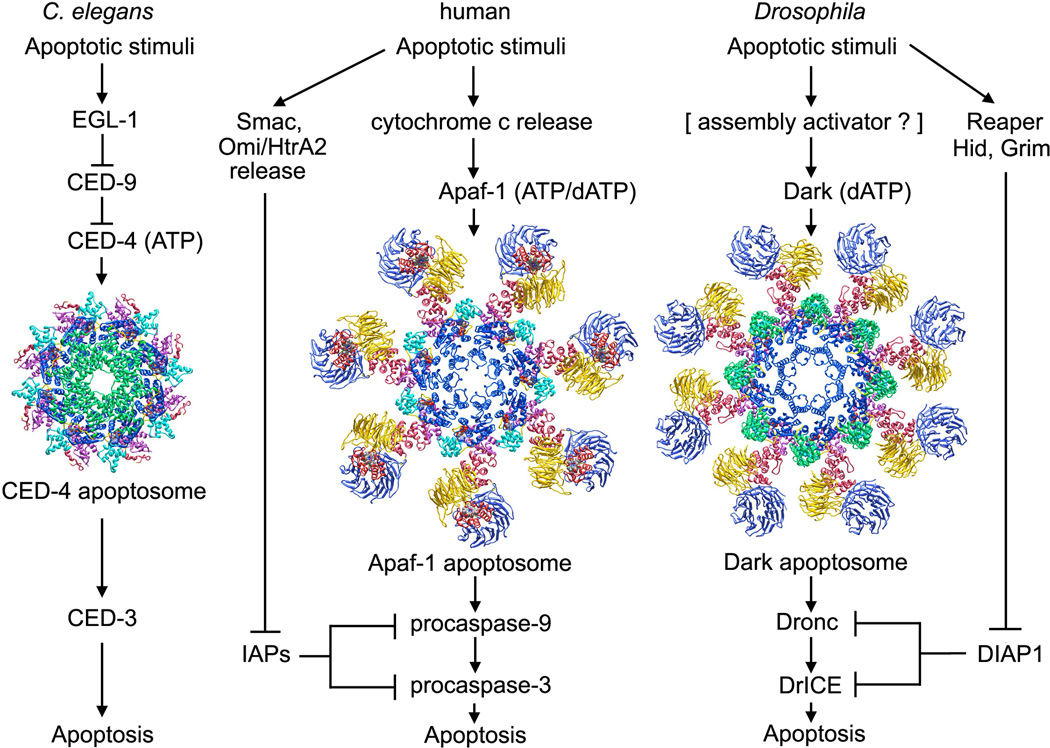
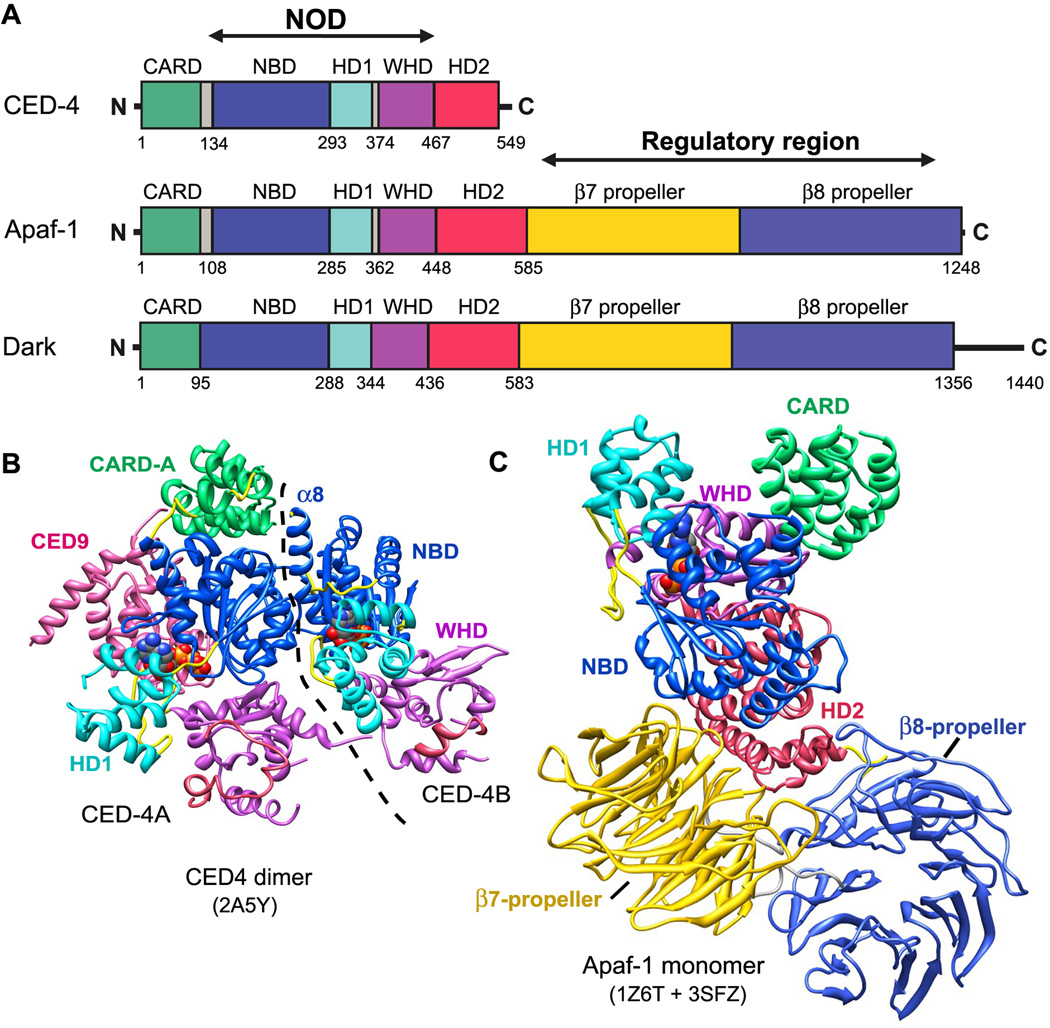
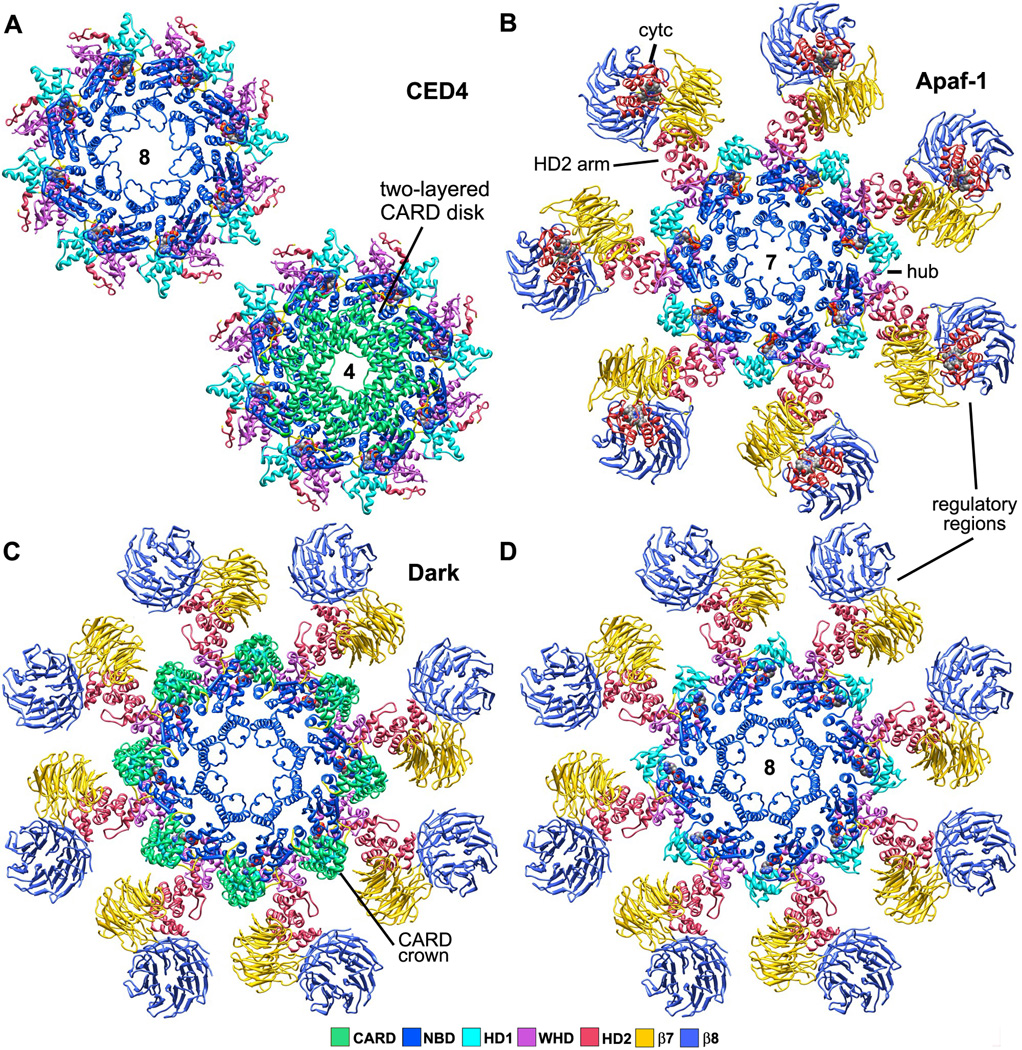
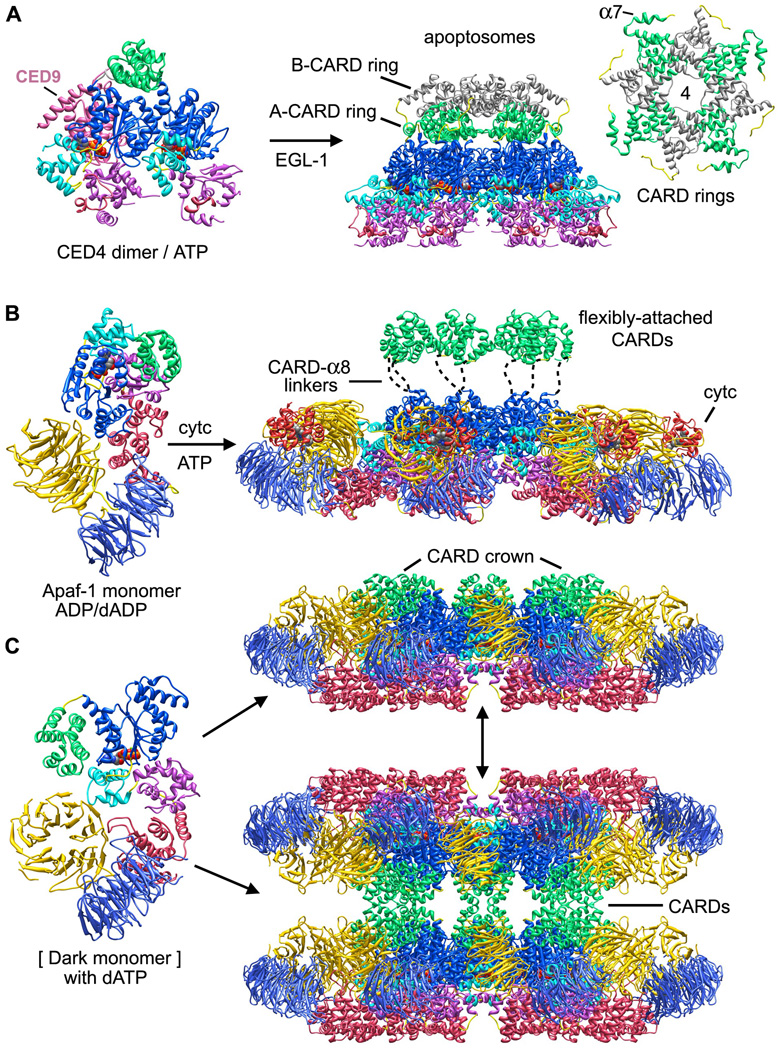
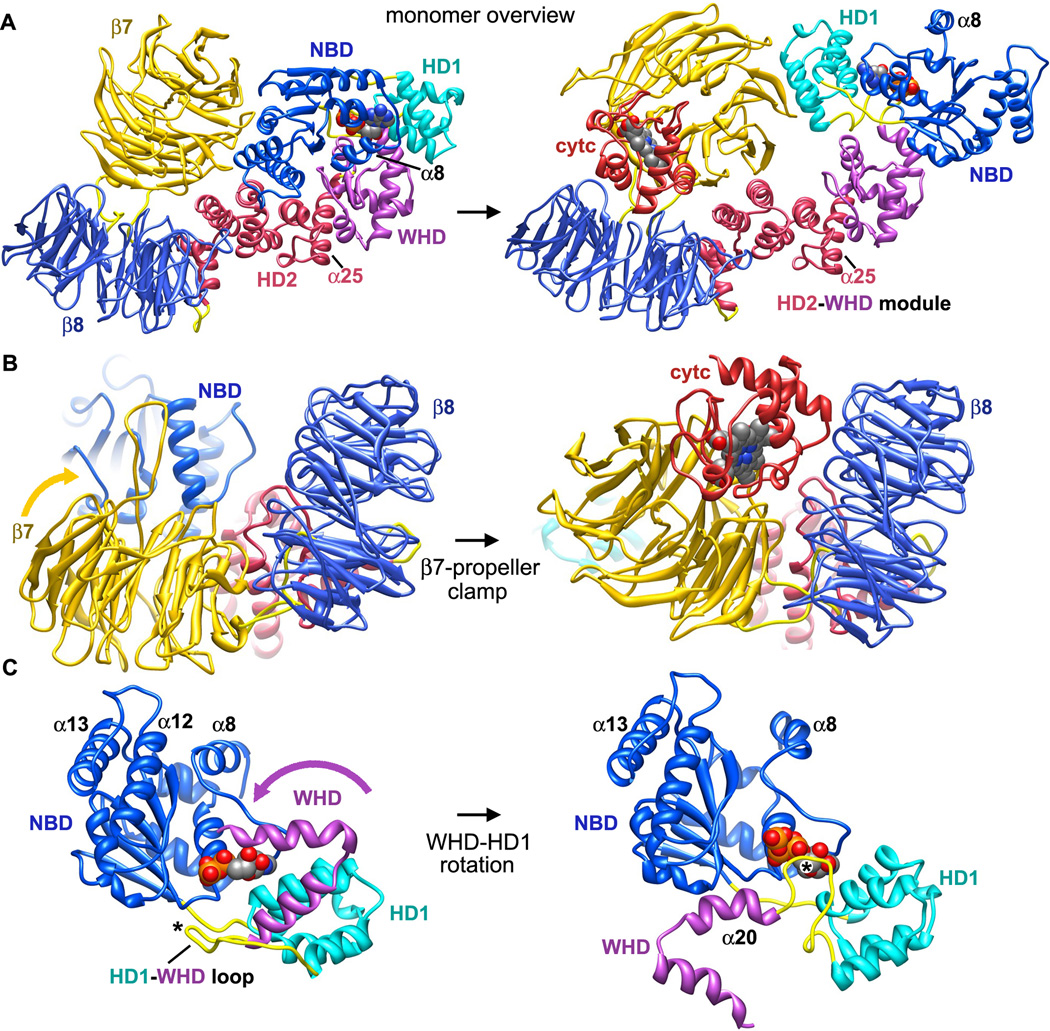
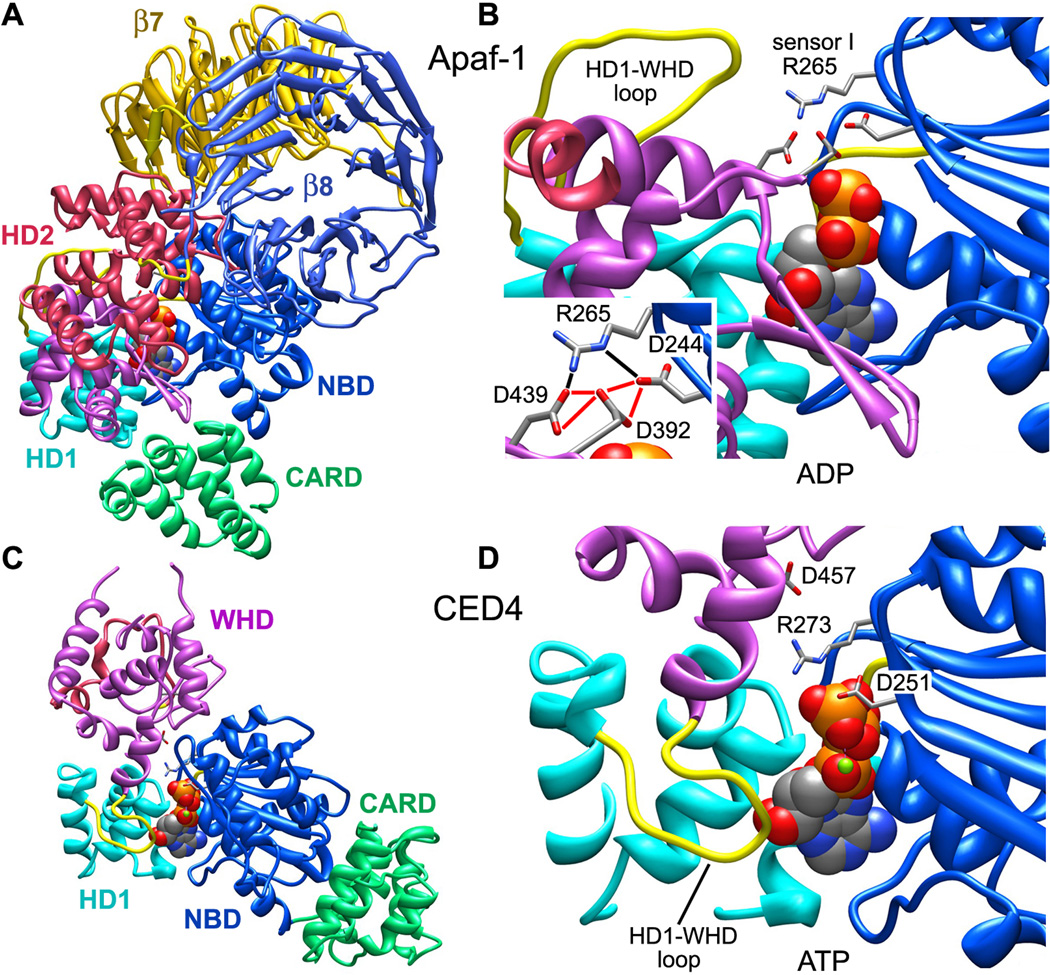
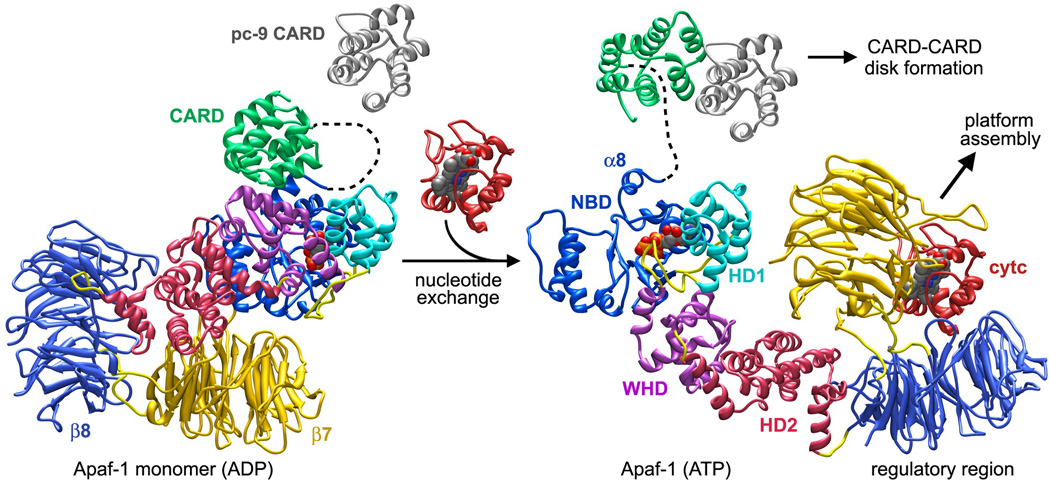
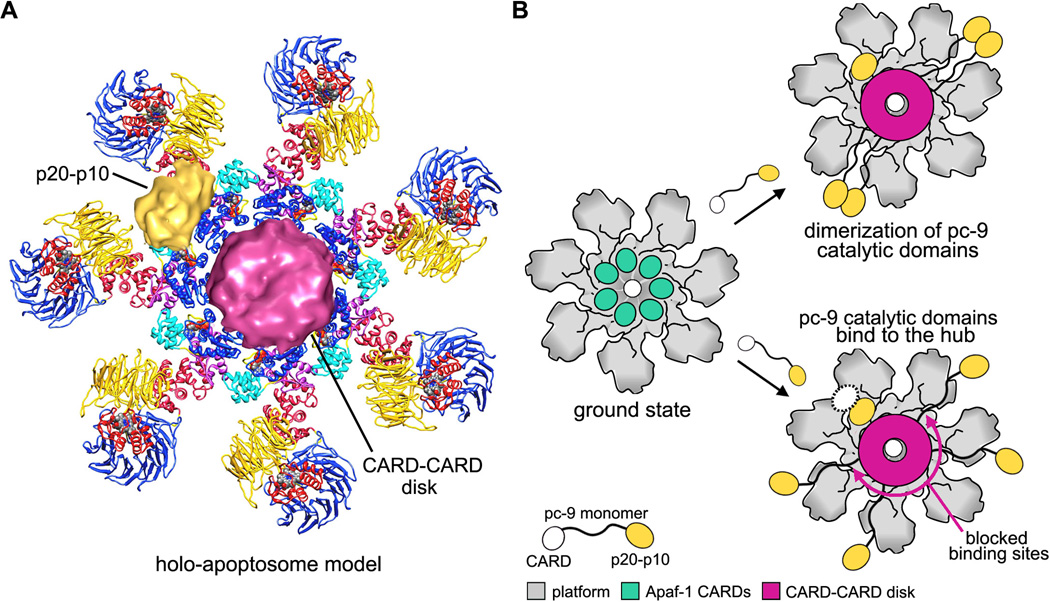
Apaf-1-like molecules assemble into a ring-like platform known as the apoptosome. This cell death platform then activates procaspases in the intrinsic cell death pathway. In this review, crystal structures of Apaf-1 monomers and CED-4 dimers have been combined with apoptosome structures to provide insights into the assembly of cell death platforms in humans, nematodes, and flies. In humans, the caspase recognition domains (CARDs) of procaspase-9 and Apaf-1 interact with each other to form a CARD-CARD disk, which interacts with the platform to create an asymmetric proteolysis machine. The disk tethers multiple pc-9 catalytic domains to the platform to raise their local concentration, and this leads to zymogen activation. These findings have now set the stage for further studies of this critical activation process on the apoptosome.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures
References
-
- Acehan D, Jiang X, Morgan DG, Heuser JE, Wang X, Akey CW. Three-dimensional structure of the apoptosome: implications for assembly, procaspase-9 binding and activation. Mol. Cell. 2002;9:423–432. - PubMed
-
- Allan LA, Morrice N, Brady S, Magee G, Pathak S, Clarke PR. Inhibition of caspase-9 through phosphorylation at Thr 125 by ERK MAPK. Nat. Cell. Biol. 2003;5(7):647–654. - PubMed
-
- Allan LA, Clarke PR. Phosphorylation of caspase-9 by CDK1/cyclin B1 protects mitotic cells against apoptosis. Mol Cell. 2007;26:301–310. - PubMed
-
- Bao Q, Lu W, Rabinowitz JD, Shi Y. Calcium blocks formation of apoptosome by preventing nucleotide exchange in Apaf-1. Mol. Cell. 2007;25:181–192. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
