

A network of high-mobility group box transcription factors programs innate interleukin-17 production
- PMID: 23562159
- PMCID: PMC3811080
- DOI: 10.1016/j.immuni.2013.01.010
A network of high-mobility group box transcription factors programs innate interleukin-17 production
Abstract
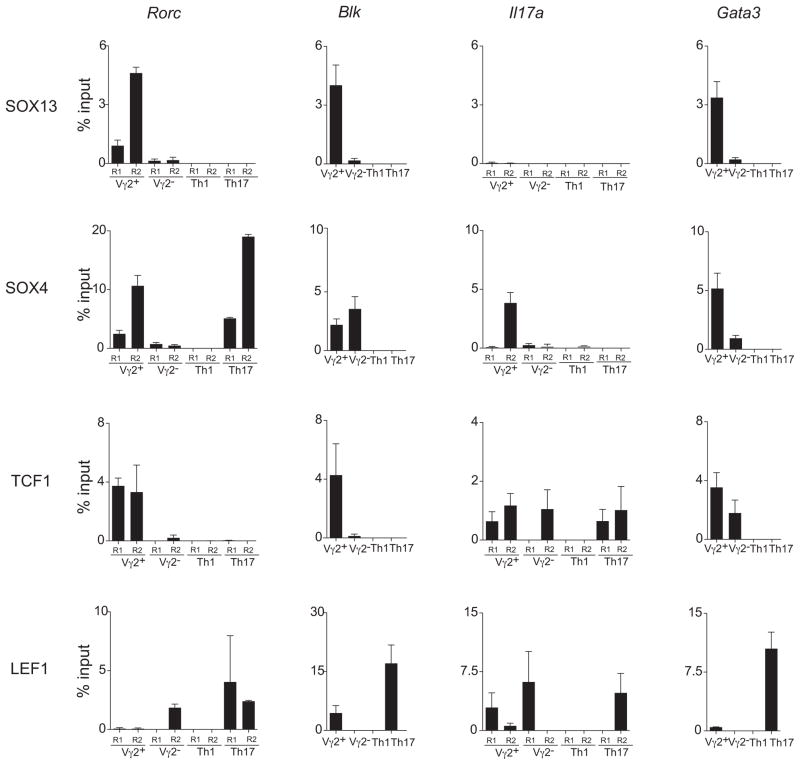
How innate lymphoid cells (ILCs) in the thymus and gut become specialized effectors is unclear. The prototypic innate-like γδ T cells (Tγδ17) are a major source of interleukin-17 (IL-17). We demonstrate that Tγδ17 cells are programmed by a gene regulatory network consisting of a quartet of high-mobility group (HMG) box transcription factors, SOX4, SOX13, TCF1, and LEF1, and not by conventional TCR signaling. SOX4 and SOX13 directly regulated the two requisite Tγδ17 cell-specific genes, Rorc and Blk, whereas TCF1 and LEF1 countered the SOX proteins and induced genes of alternate effector subsets. The T cell lineage specification factor TCF1 was also indispensable for the generation of IL-22 producing gut NKp46(+) ILCs and restrained cytokine production by lymphoid tissue inducer-like effectors. These results indicate that similar gene network architecture programs innate sources of IL-17, independent of anatomical origins.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures





References
-
- Brustle A, Heink S, Huber M, Rosenplanter C, Stadelmann C, Yu P, Arpaia E, Mak TW, Kamradt T, Lohoff M. The development of inflammatory Th17 cells requires interferon-regulatory factor 4. Nat Immunol. 2007;8:958–966. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 AR054153/AR/NIAMS NIH HHS/United States
- UL1 TR000439/TR/NCATS NIH HHS/United States
- R01 AI101048/AI/NIAID NIH HHS/United States
- R01 CA100382/CA/NCI NIH HHS/United States
- T32 AR007465/AR/NIAMS NIH HHS/United States
- R01 AI101301/AI/NIAID NIH HHS/United States
- AR54153/AR/NIAMS NIH HHS/United States
- CA100382/CA/NCI NIH HHS/United States
- P30 DK026743/DK/NIDDK NIH HHS/United States
- K08 AR061437/AR/NIAMS NIH HHS/United States
- AI072073/AI/NIAID NIH HHS/United States
- AI084987/AI/NIAID NIH HHS/United States
- R01 AI084987/AI/NIAID NIH HHS/United States
- R24 AI072073/AI/NIAID NIH HHS/United States
- T32 AI007349/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
