

Different thresholds of Wnt-Frizzled 7 signaling coordinate proliferation, morphogenesis and fate of endoderm progenitor cells
- PMID: 23562607
- PMCID: PMC4263287
- DOI: 10.1016/j.ydbio.2013.02.024
Different thresholds of Wnt-Frizzled 7 signaling coordinate proliferation, morphogenesis and fate of endoderm progenitor cells
Abstract
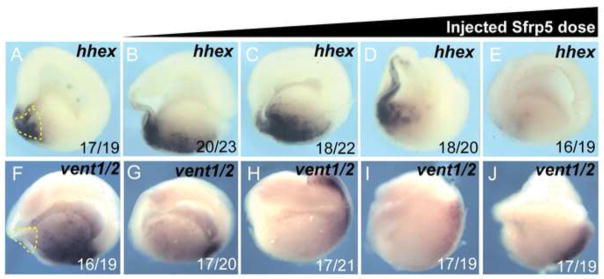
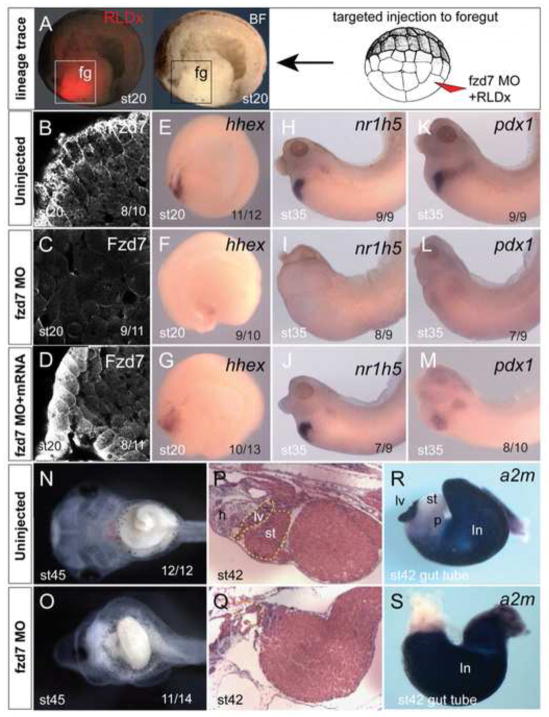
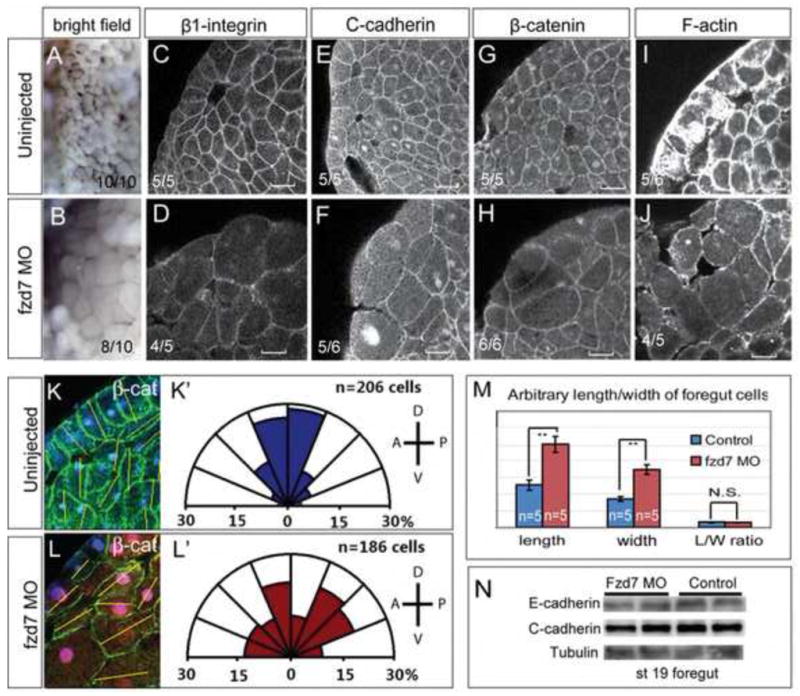
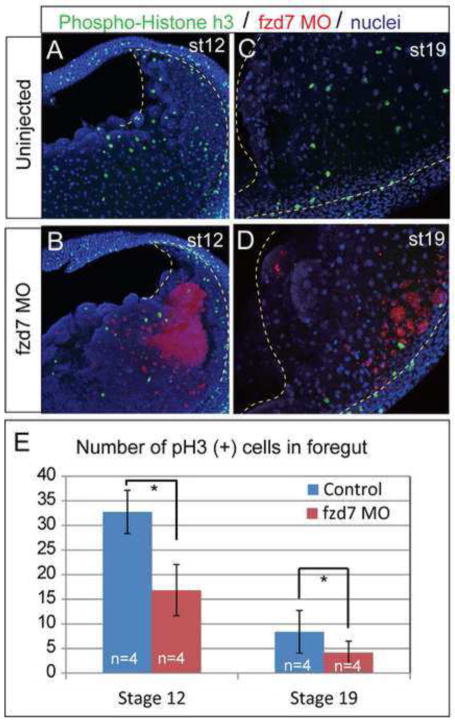
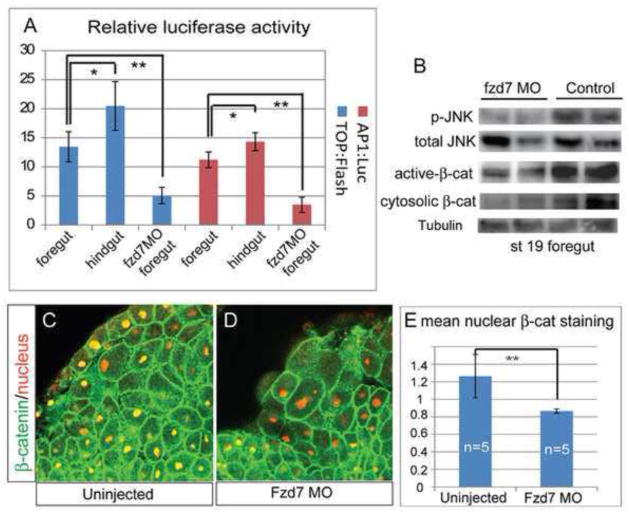
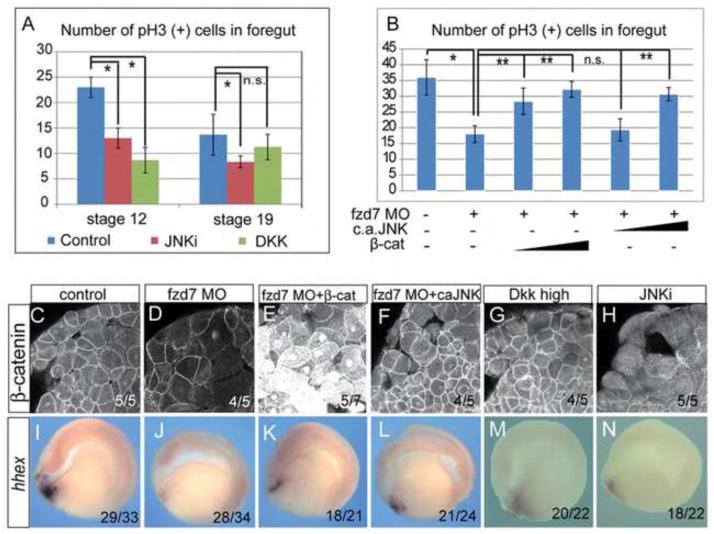
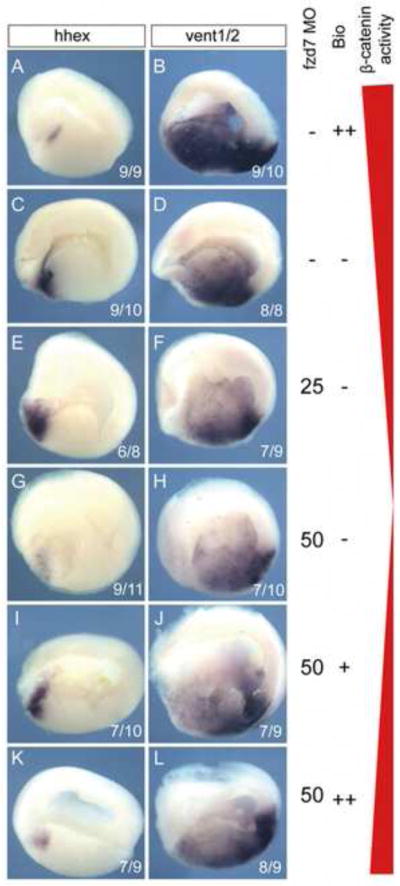
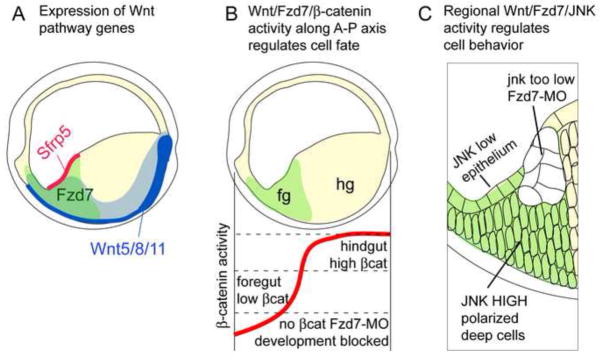
Wnt signaling has multiple dynamic roles during development of the gastrointestinal and respiratory systems. Differential Wnt signaling is thought to be a critical step in Xenopus endoderm patterning such that during late gastrula and early somite stages of embryogenesis, Wnt activity must be suppressed in the anterior to allow the specification of foregut progenitors. However, the foregut endoderm also expresses the Wnt-receptor Frizzled 7 (Fzd7) as well as several Wnt ligands suggesting that the current model may be too simple. In this study, we show that Fzd7 is required to transduce a low level of Wnt signaling that is essential to maintain foregut progenitors. Foregut-specific Fzd7-depletion from the Xenopus foregut resulted in liver and pancreas agenesis. Fzd7-depleted embryos failed to maintain the foregut progenitor marker hhex and exhibited decreased proliferation; in addition the foregut cells were enlarged with a randomized orientation. We show that in the foregut Fzd7 signals via both the Wnt/β-catenin and Wnt/JNK pathways and that different thresholds of Wnt-Fzd7 activity coordinate progenitor cell fate, proliferation and morphogenesis.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Syndecan4 coordinates Wnt/JNK and BMP signaling to regulate foregut progenitor development.Dev Biol. 2016 Aug 1;416(1):187-199. doi: 10.1016/j.ydbio.2016.05.025. Epub 2016 May 24. Dev Biol. 2016. PMID: 27235146 Free PMC article.
-
Wnt2/2b and beta-catenin signaling are necessary and sufficient to specify lung progenitors in the foregut.Dev Cell. 2009 Aug;17(2):290-8. doi: 10.1016/j.devcel.2009.06.005. Dev Cell. 2009. PMID: 19686689 Free PMC article.
-
Repression of Wnt/beta-catenin signaling in the anterior endoderm is essential for liver and pancreas development.Development. 2007 Jun;134(12):2207-17. doi: 10.1242/dev.001230. Epub 2007 May 16. Development. 2007. PMID: 17507400
-
Beta-catenin signaling in hepatic development and progenitors: which way does the WNT blow?Dev Dyn. 2011 Mar;240(3):486-500. doi: 10.1002/dvdy.22522. Epub 2010 Dec 23. Dev Dyn. 2011. PMID: 21337461 Free PMC article. Review.
-
Heads or tails? Amphioxus and the evolution of anterior-posterior patterning in deuterostomes.Dev Biol. 2002 Jan 15;241(2):209-28. doi: 10.1006/dbio.2001.0503. Dev Biol. 2002. PMID: 11784106 Review.
Cited by
-
Circular RNA circCSPP1 knockdown attenuates doxorubicin resistance and suppresses tumor progression of colorectal cancer via miR-944/FZD7 axis.Cancer Cell Int. 2021 Mar 4;21(1):153. doi: 10.1186/s12935-021-01855-6. Cancer Cell Int. 2021. PMID: 33663510 Free PMC article.
-
Orchestrating liver development.Development. 2015 Jun 15;142(12):2094-108. doi: 10.1242/dev.114215. Development. 2015. PMID: 26081571 Free PMC article. Review.
-
Frogs as integrative models for understanding digestive organ development and evolution.Semin Cell Dev Biol. 2016 Mar;51:92-105. doi: 10.1016/j.semcdb.2016.02.001. Epub 2016 Feb 3. Semin Cell Dev Biol. 2016. PMID: 26851628 Free PMC article. Review.
-
ATP4 and ciliation in the neuroectoderm and endoderm of Xenopus embryos and tadpoles.Data Brief. 2015 Apr 20;4:22-31. doi: 10.1016/j.dib.2015.04.003. eCollection 2015 Sep. Data Brief. 2015. PMID: 26217756 Free PMC article.
-
Genomic integration of Wnt/β-catenin and BMP/Smad1 signaling coordinates foregut and hindgut transcriptional programs.Development. 2017 Apr 1;144(7):1283-1295. doi: 10.1242/dev.145789. Epub 2017 Feb 20. Development. 2017. PMID: 28219948 Free PMC article.
References
-
- Bovolenta P, Rodriguez J, Esteve P. Frizzled/RYK mediated signalling in axon guidance. Development. 2006;133:4399–4408. - PubMed
-
- Brown JD, Hallagan SE, McGrew LL, Miller JR, Moon RT. The maternal Xenopus beta-catenin signaling pathway, activated by frizzled homologs, induces goosecoid in a cell non-autonomous manner. Development, growth & differentiation. 2000;42:347–357. - PubMed
-
- Cha SW, Tadjuidje E, Tao Q, Wylie C, Heasman J. Wnt5a and Wnt11 interact in a maternal Dkk1-regulated fashion to activate both canonical and non-canonical signaling in Xenopus axis formation. Development. 2008;135:3719–3729. - PubMed
-
- Cha SW, Tadjuidje E, White J, Wells J, Mayhew C, Wylie C, Heasman J. Wnt11/5a complex formation caused by tyrosine sulfation increases canonical signaling activity. Curr Biol. 2009;19:1573–1580. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
