

The bacterial DnaC helicase loader is a DnaB ring breaker
- PMID: 23562643
- PMCID: PMC3670212
- DOI: 10.1016/j.cell.2013.03.006
The bacterial DnaC helicase loader is a DnaB ring breaker
Abstract
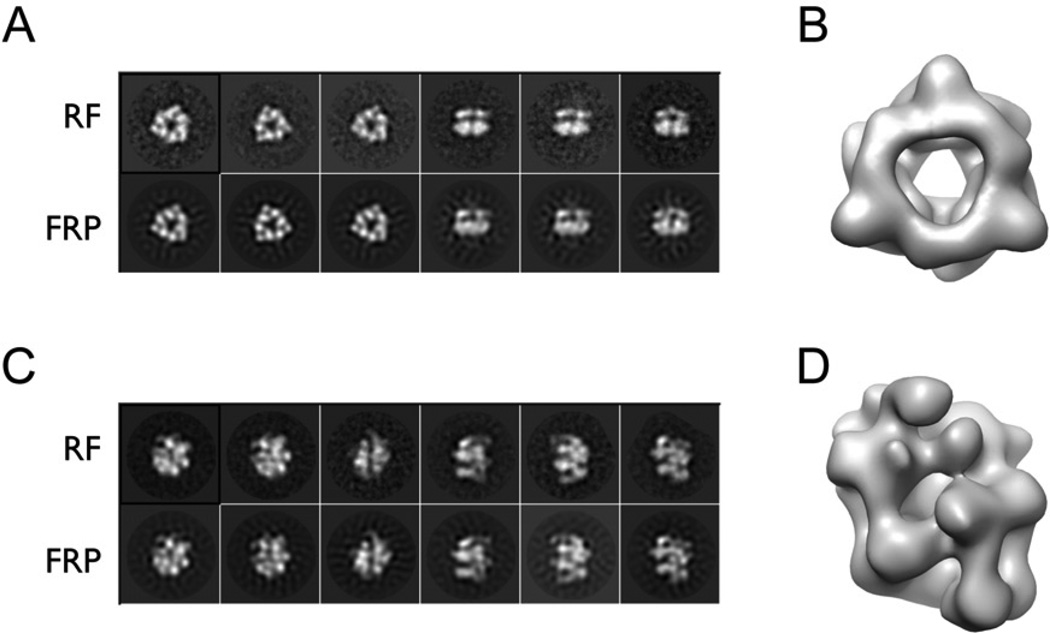
Dedicated AAA+ ATPases deposit hexameric ring-shaped helicases onto DNA to promote replication in cellular organisms. To understand how loading occurs, we used electron microscopy and small angle X-ray scattering (SAXS) to determine the ATP-bound structure of the intact E. coli DnaB⋅DnaC helicase/loader complex. The 480 kDa dodecamer forms a three-tiered assembly, in which DnaC adopts a spiral configuration that remodels N-terminal scaffolding and C-terminal motor regions of DnaB to produce a clear break in the helicase ring. Surprisingly, DnaC's AAA+ fold is dispensable for ring remodeling because the DnaC isolated helicase-binding domain can both load DnaB onto DNA and increase the efficiency by which the helicase acts on substrates in vitro. Our data demonstrate that DnaC opens DnaB by a mechanism akin to that of polymerase clamp loaders and indicate that bacterial replicative helicases, like their eukaryotic counterparts, possess autoregulatory elements that influence how hexameric motor domains are loaded onto and unwind DNA.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures






References
-
- Ason B, Bertram JG, Hingorani MM, Beechem JM, O’Donnell M, Goodman MF, Bloom LB. A model for Escherichia coli DNA polymerase III holoenzyme assembly at primer/template ends. DNA triggers a change in binding specificity of the gamma complex clamp loader. J Biol Chem. 2000;275:3006–3015. - PubMed
-
- Bailey S, Eliason WK, Steitz TA. Structure of hexameric DnaB helicase and its complex with a domain of DnaG primase. Science. 2007b;318:459–463. - PubMed
-
- Baker TA, Bell SP. Polymerases and the replisome: machines within machines. Cell. 1998;92:295–305. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
