

Non-cell-autonomous tumor suppression by p53
- PMID: 23562644
- PMCID: PMC3702034
- DOI: 10.1016/j.cell.2013.03.020
Non-cell-autonomous tumor suppression by p53
Abstract
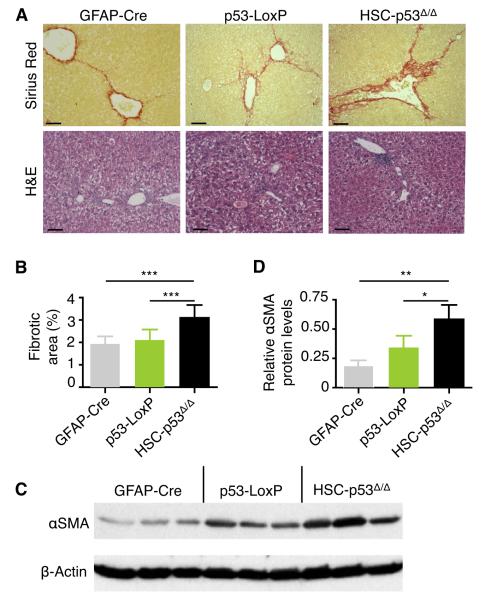
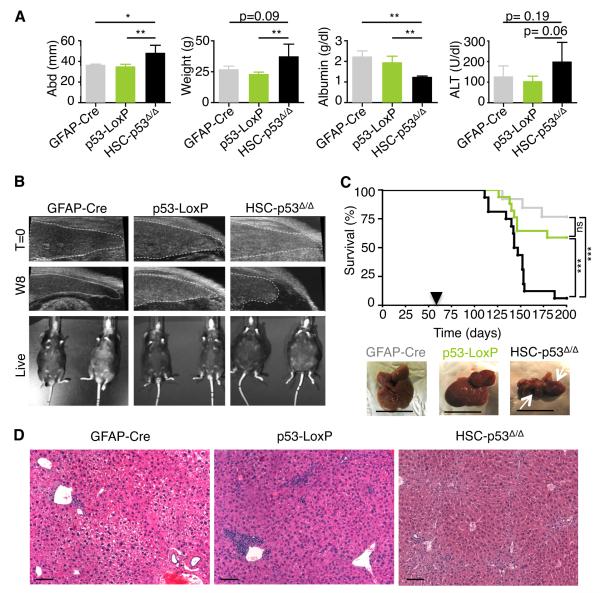
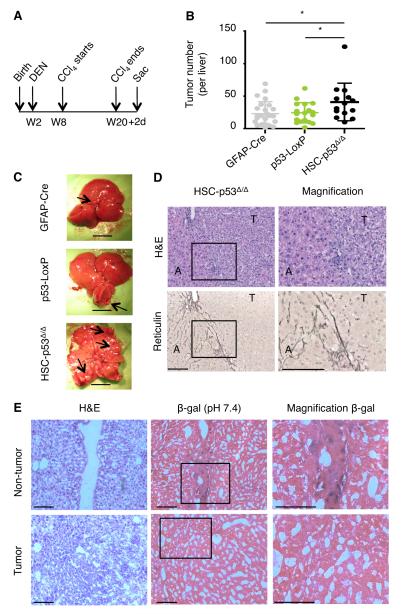
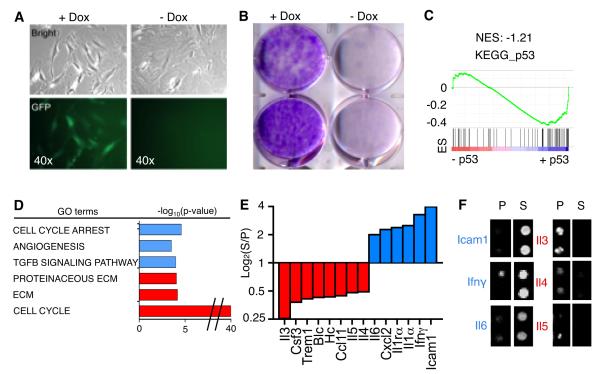
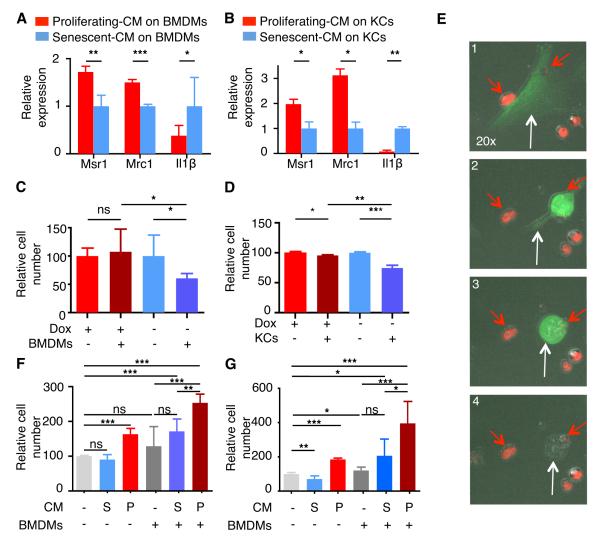
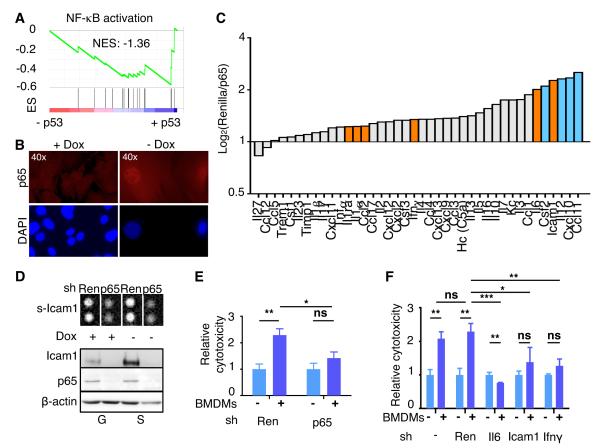
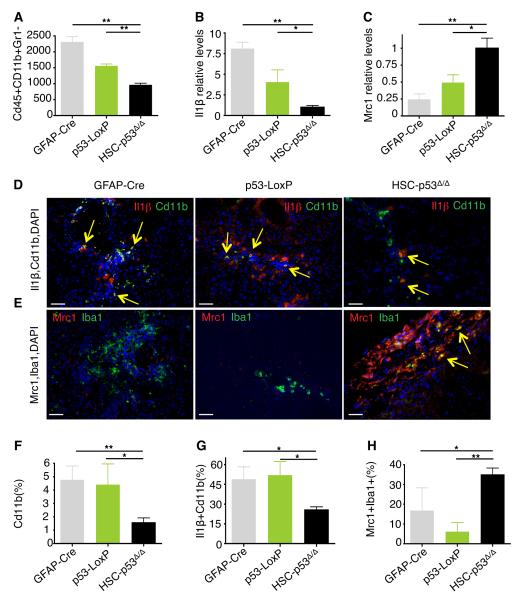
The p53 tumor suppressor can restrict malignant transformation by triggering cell-autonomous programs of cell-cycle arrest or apoptosis. p53 also promotes cellular senescence, a tumor-suppressive program that involves stable cell-cycle arrest and secretion of factors that modify the tissue microenvironment. In the presence of chronic liver damage, we show that ablation of a p53-dependent senescence program in hepatic stellate cells increases liver fibrosis and cirrhosis associated with reduced survival and enhances the transformation of adjacent epithelial cells into hepatocellular carcinoma. p53-expressing senescent stellate cells release factors that skew macrophage polarization toward a tumor-inhibiting M1-state capable of attacking senescent cells in culture, whereas proliferating p53-deficient stellate cells secrete factors that stimulate polarization of macrophages into a tumor-promoting M2-state and enhance the proliferation of premalignant cells. Hence, p53 can act non-cell autonomously to suppress tumorigenesis by promoting an antitumor microenvironment, in part, through secreted factors that modulate macrophage function.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Tumor suppression: p53 alters immune surveillance to restrain liver cancer.Curr Biol. 2013 Jun 17;23(12):R527-30. doi: 10.1016/j.cub.2013.04.076. Curr Biol. 2013. PMID: 23787049 Free PMC article.
References
-
- Acosta JC, O’Loghlen A, Banito A, Guijarro MV, Augert A, Raguz S, Fumagalli M, Da Costa M, Brown C, Popov N, et al. Chemokine signaling via the CXCR2 receptor reinforces senescence. Cell. 2008;133:1006–1018. - PubMed
-
- Biswas SK, Mantovani A. Macrophage plasticity and interaction with lymphocyte subsets: cancer as a paradigm. Nat Immunol. 2010;11:889–896. - PubMed
-
- Campisi J, d’Adda di Fagagna F. Cellular senescence: when bad things happen to good cells. Nat Rev Mol Cell Biol. 2007;8:729–740. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
