

Protein sorting, targeting and trafficking in photoreceptor cells
- PMID: 23562855
- PMCID: PMC3759535
- DOI: 10.1016/j.preteyeres.2013.03.002
Protein sorting, targeting and trafficking in photoreceptor cells
Abstract
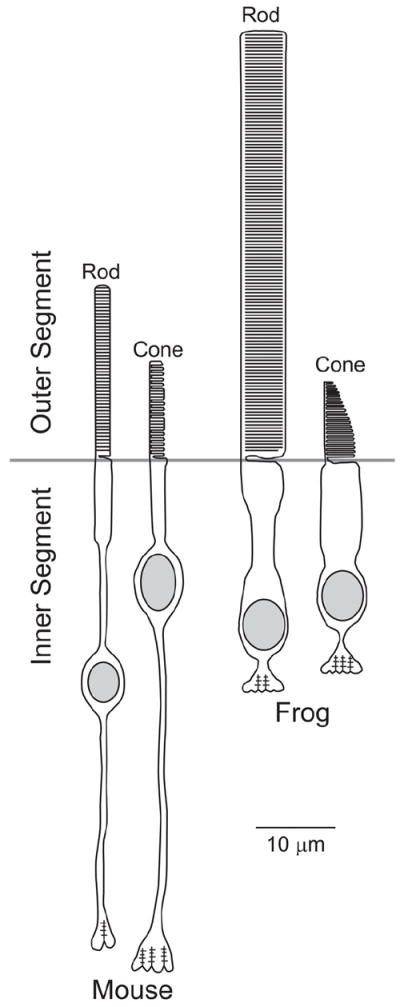
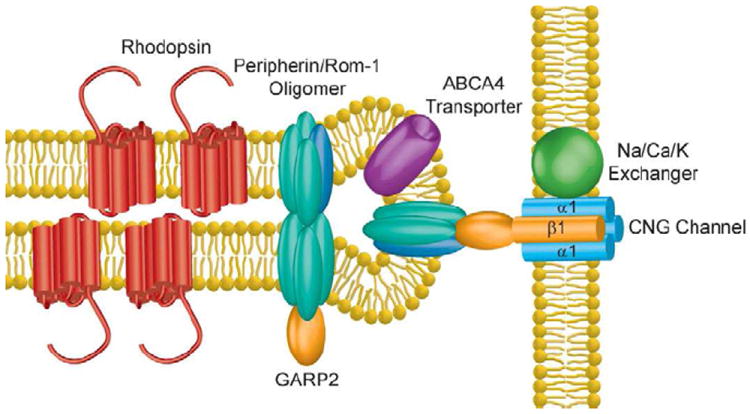
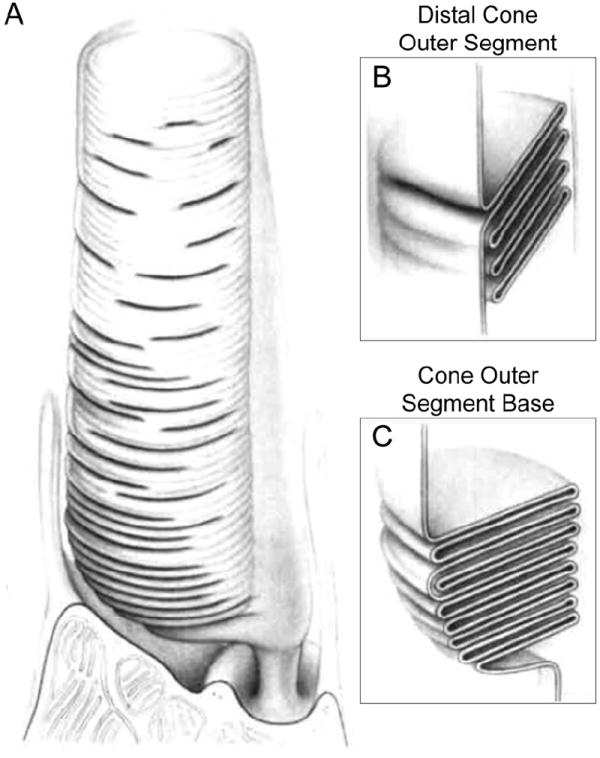
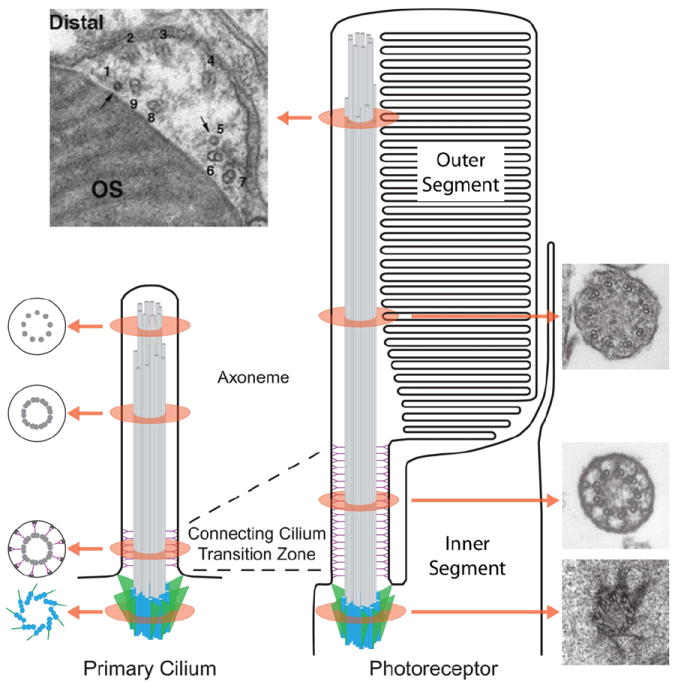
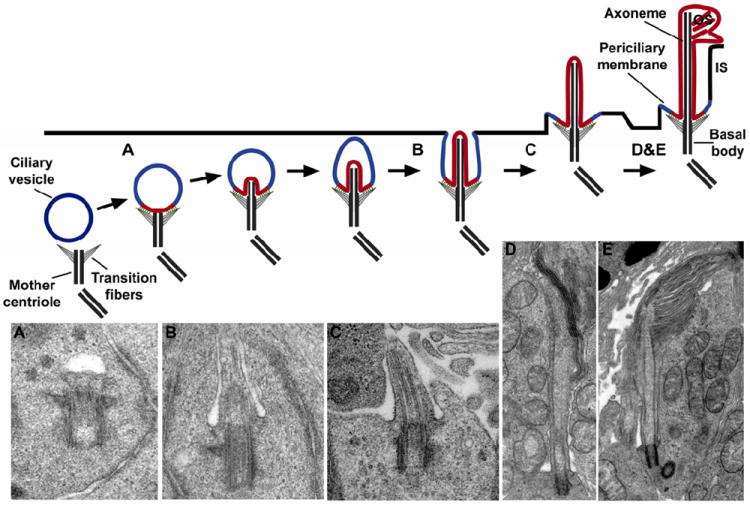
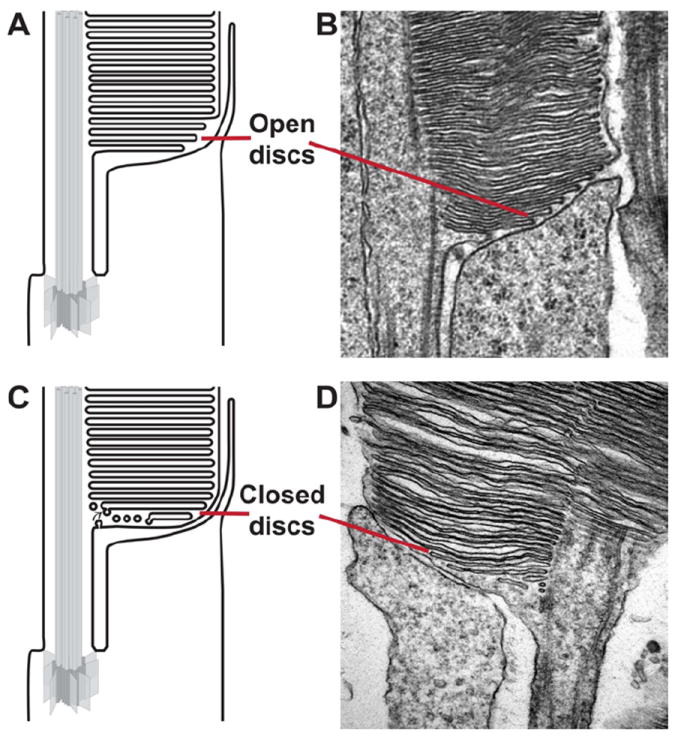
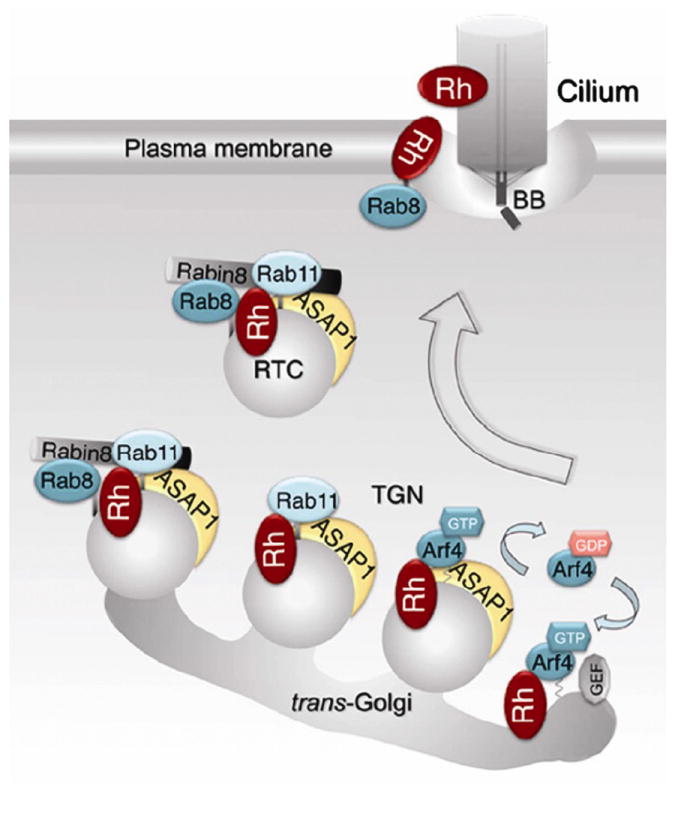
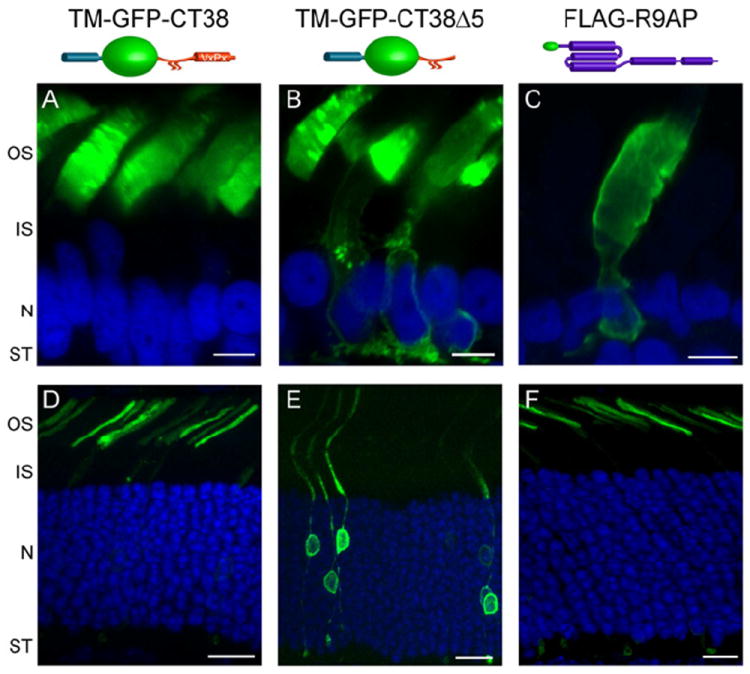
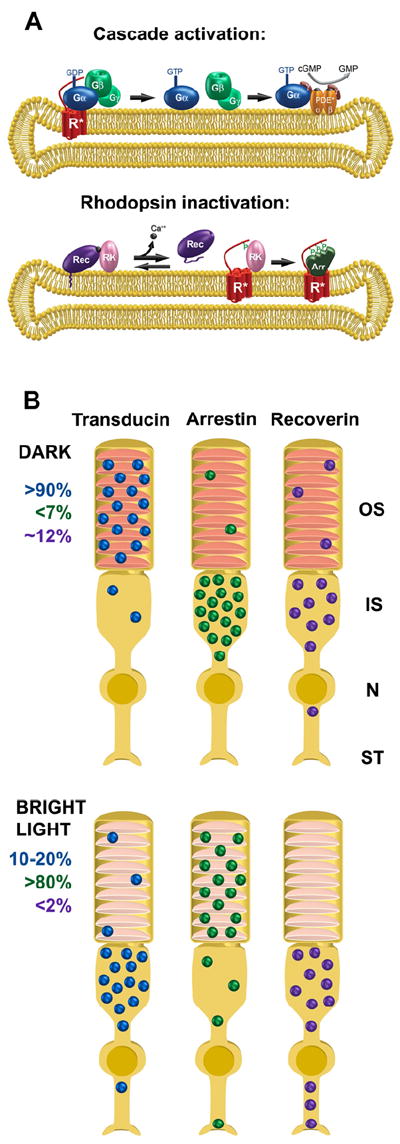
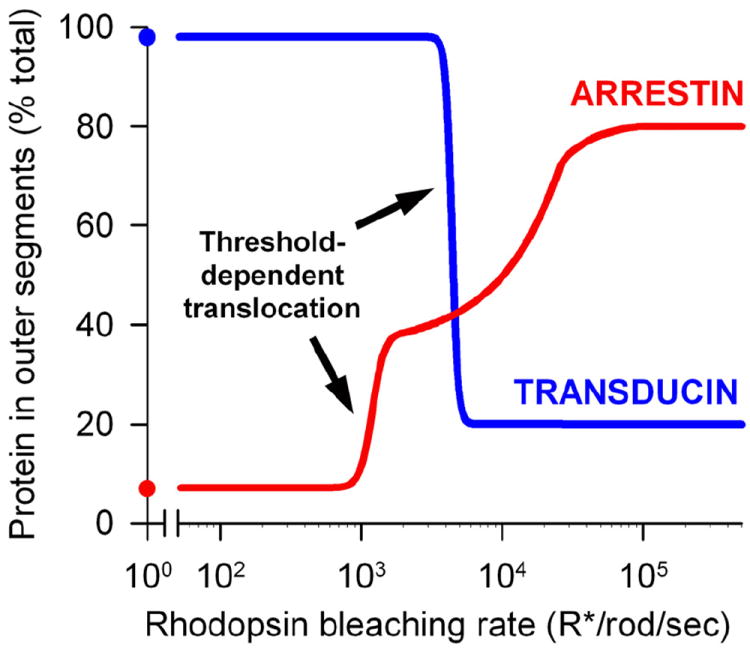
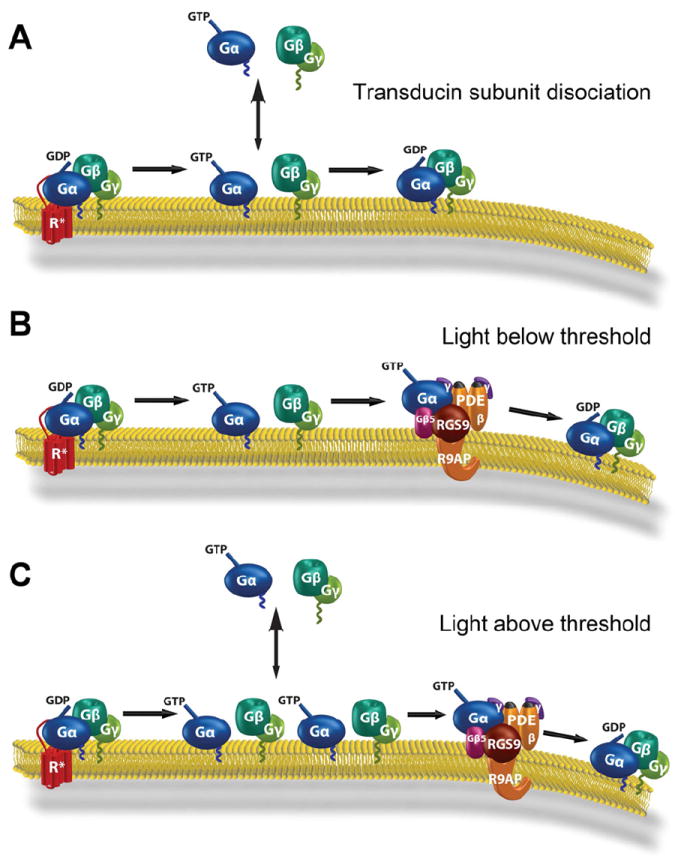
Vision is the most fundamental of our senses initiated when photons are absorbed by the rod and cone photoreceptor neurons of the retina. At the distal end of each photoreceptor resides a light-sensing organelle, called the outer segment, which is a modified primary cilium highly enriched with proteins involved in visual signal transduction. At the proximal end, each photoreceptor has a synaptic terminal, which connects this cell to the downstream neurons for further processing of the visual information. Understanding the mechanisms involved in creating and maintaining functional compartmentalization of photoreceptor cells remains among the most fascinating topics in ocular cell biology. This review will discuss how photoreceptor compartmentalization is supported by protein sorting, targeting and trafficking, with an emphasis on the best-studied cases of outer segment-resident proteins.
Keywords: Cilium; Outer segment; Photoreceptor; Protein targeting; Protein trafficking.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures
References
-
- Adler AJ, Klucznik KM. Proteins and glycoproteins of the bovine interphotoreceptor matrix: composition and fractionation. Exp Eye Res. 1982;34:423–434. - PubMed
-
- Adler AJ, Severin KM. Proteins of the bovine interphotoreceptor matrix: tissues of origin. Exp Eye Res. 1981;32:755–769. - PubMed
-
- Anant JS, Fung BK. In vivo farnesylation of rat rhodopsin kinase. Biochem Biophys Res Commun. 1992;183:468–473. - PubMed
-
- Anant JS, Ong OC, Xie HY, Clarke S, O’Brien PJ, Fung BK. In vivo differential prenylation of retinal cyclic GMP phosphodiesterase catalytic subunits. J Biol Chem. 1992;267:687–690. - PubMed
-
- Anderson DH, Fisher SK. Disc shedding in rodlike and conelike photoreceptors of tree squirrels. Science. 1975;187:953–955. - PubMed
Publication types
MeSH terms
Grants and funding
- P30 EY005722/EY/NEI NIH HHS/United States
- T32 GM007184/GM/NIGMS NIH HHS/United States
- EY020542/EY/NEI NIH HHS/United States
- F32 EY022508/EY/NEI NIH HHS/United States
- F31 EY022862/EY/NEI NIH HHS/United States
- EY022862/EY/NEI NIH HHS/United States
- EY022508/EY/NEI NIH HHS/United States
- EY010336/EY/NEI NIH HHS/United States
- R01 EY012859/EY/NEI NIH HHS/United States
- R01 EY010336/EY/NEI NIH HHS/United States
- EY012859/EY/NEI NIH HHS/United States
- EY05722/EY/NEI NIH HHS/United States
- R01 EY020542/EY/NEI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
