

Promoter sequence determines the relationship between expression level and noise
- PMID: 23565060
- PMCID: PMC3614515
- DOI: 10.1371/journal.pbio.1001528
Promoter sequence determines the relationship between expression level and noise
Abstract
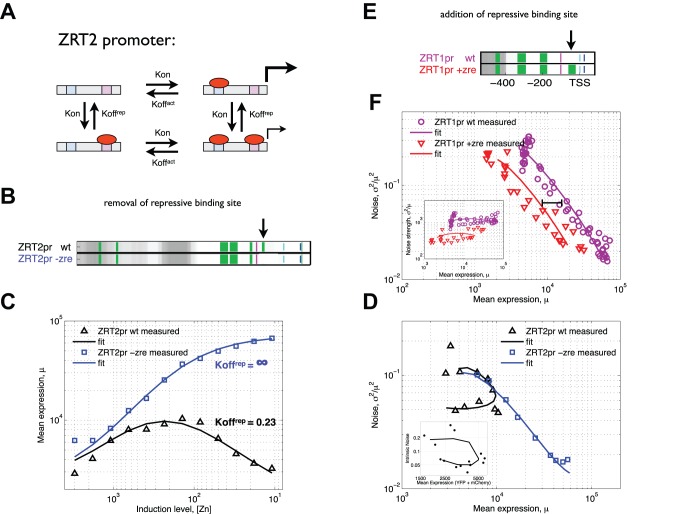
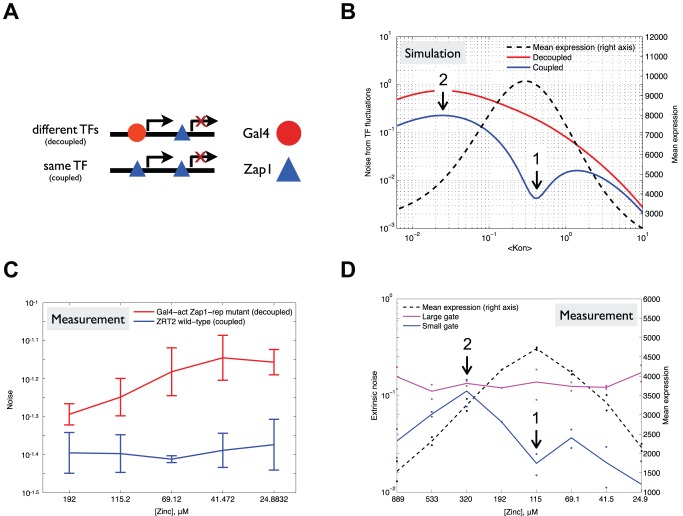
The ability of cells to accurately control gene expression levels in response to extracellular cues is limited by the inherently stochastic nature of transcriptional regulation. A change in transcription factor (TF) activity results in changes in the expression of its targets, but the way in which cell-to-cell variability in expression (noise) changes as a function of TF activity, and whether targets of the same TF behave similarly, is not known. Here, we measure expression and noise as a function of TF activity for 16 native targets of the transcription factor Zap1 that are regulated by it through diverse mechanisms. For most activated and repressed Zap1 targets, noise decreases as expression increases. Kinetic modeling suggests that this is due to two distinct Zap1-mediated mechanisms that both change the frequency of transcriptional bursts. Notably, we found that another mechanism of repression by Zap1, which is encoded in the promoter DNA, likely decreases the size of transcriptional bursts, producing a unique transcriptional state characterized by low expression and low noise. In addition, we find that further reduction in noise is achieved when a single TF both activates and represses a single target gene. Our results suggest a global principle whereby at low TF concentrations, the dominant source of differences in expression between promoters stems from differences in burst frequency, whereas at high TF concentrations differences in burst size dominate. Taken together, we show that the precise amount by which noise changes with expression is specific to the regulatory mechanism of transcription and translation that acts at each gene.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





References
-
- Benzer S (1953) Induced synthesis of enzymes in bacteria analyzed at the cellular level. Biochim Biophys Acta 11: 383–395. - PubMed
-
- Newman JRS, Ghaemmaghami S, Ihmels J, Breslow DK, Noble M, et al. (2006) Single-cell proteomic analysis of S. cerevisiae reveals the architecture of biological noise. Nature 441: 840–846 doi:10.1038/nature04785 - DOI - PubMed
-
- Bar-Even A, Paulsson J, Maheshri N, Carmi M, O'Shea E, et al. (2006) Noise in protein expression scales with natural protein abundance. Nat Genet 38: 636–643 doi:10.1038/ng1807 - DOI - PubMed
-
- Taniguchi Y, Choi PJ, Li GW, Chen H, Babu M, et al. (2010) Quantifying E. coli proteome and transcriptome with single-molecule sensitivity in single cells. Science 329: 533–538 doi:10.1126/science.1188308 - DOI - PMC - PubMed
-
- Maheshri N, O'Shea EK (2007) Living with noisy genes: how cells function reliably with inherent variability in gene expression. Annu Rev Biophys Biomol Struct 36: 413–434 doi:10.1146/annurev.biophys.36.040306.132705 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
