

P2X7 integrates PI3K/AKT and AMPK-PRAS40-mTOR signaling pathways to mediate tumor cell death
- PMID: 23565201
- PMCID: PMC3615040
- DOI: 10.1371/journal.pone.0060184
P2X7 integrates PI3K/AKT and AMPK-PRAS40-mTOR signaling pathways to mediate tumor cell death
Abstract
Background: Extracellular adenosine triphosphate (ATP) functions as a novel danger signal that boosts antitumor immunity and can also directly kill tumor cells. We have previously reported that chronic exposure of tumor cells to ATP provokes P2X7-mediated tumor cell death, by as yet incompletely defined molecular mechanisms.
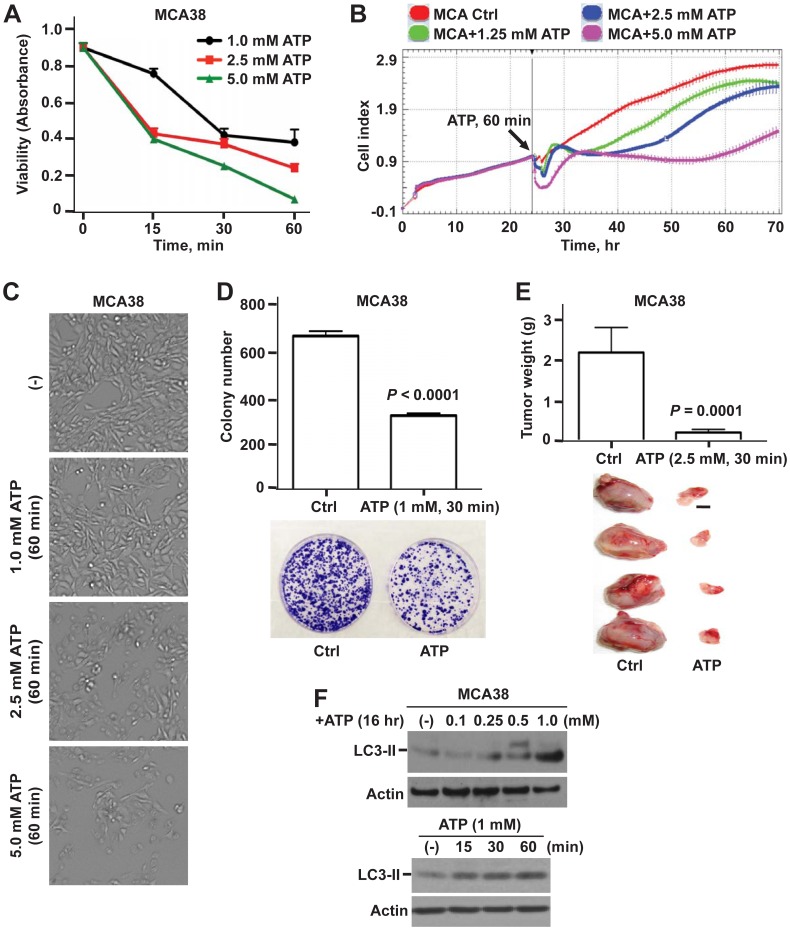
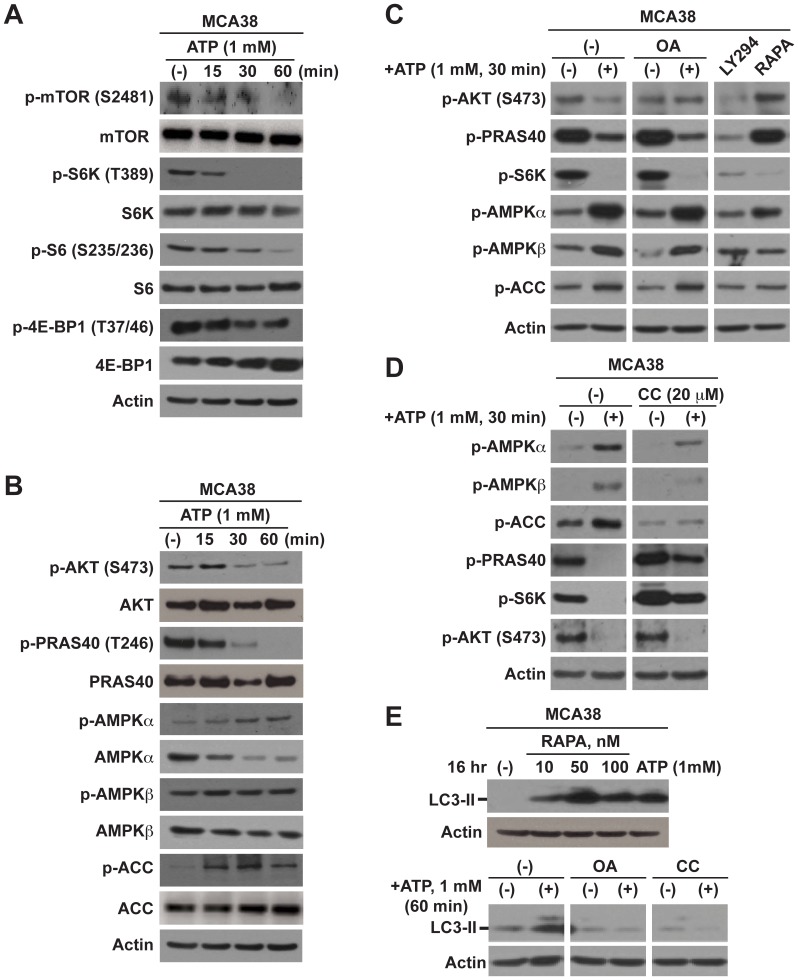
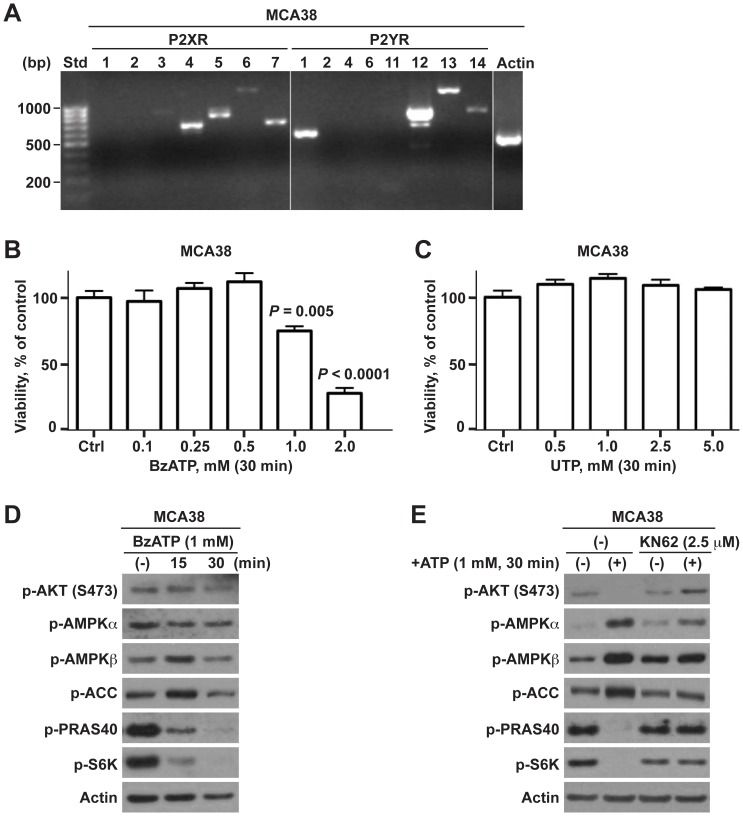
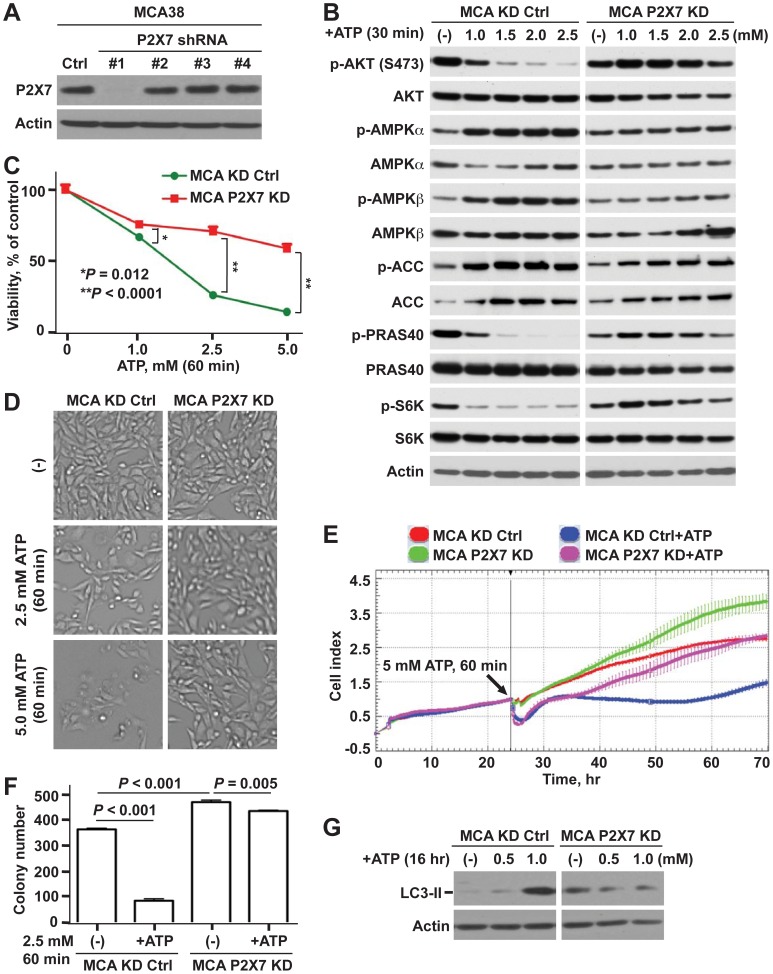
Methodology/principal findings: Here, we show that acute exposure of tumor cells to ATP results in rapid cytotoxic effects impacting several aspects of cell growth/survival, leading to inhibition of tumor growth in vitro and in vivo. Using agonist and antagonist studies together with generation of P2X7 deficient tumor cell lines by lentiviral shRNA delivery system, we confirm P2X7 to be the central control node transmitting extracellular ATP signals. We identify that downstream intracellular signaling regulatory networks implicate two signaling pathways: the known P2X7-PI3K/AKT axis and remarkably a novel P2X7-AMPK-PRAS40-mTOR axis. When exposed to high levels of extracellular ATP, these two signaling axes perturb the balance between growth and autophagy, thereby promoting tumor cell death.
Conclusions: Our study defines novel molecular mechanisms underpinning the antitumor actions of P2X7 and provides a further rationale for purine-based drugs in targeted cancer therapy.
Conflict of interest statement
Figures
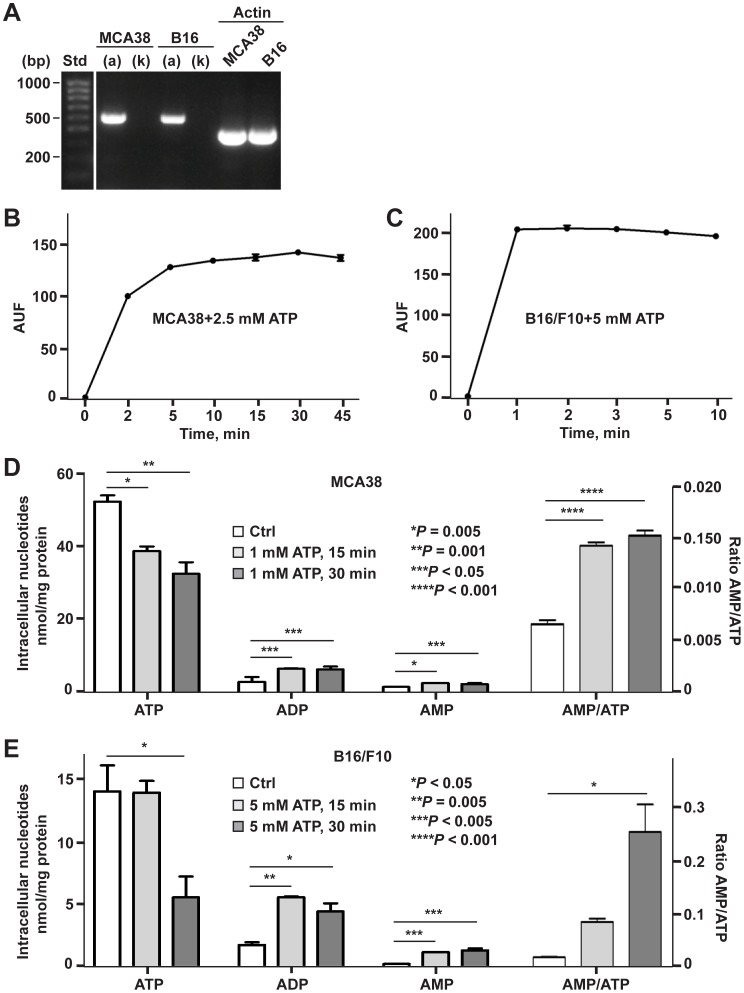
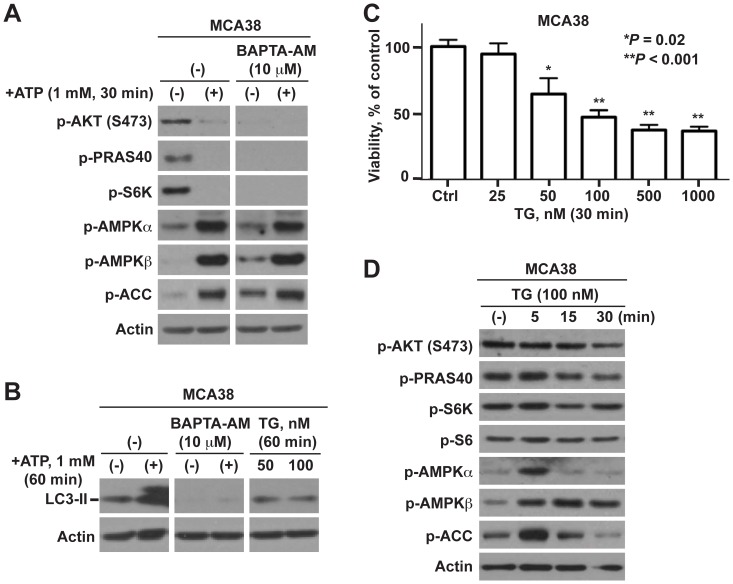
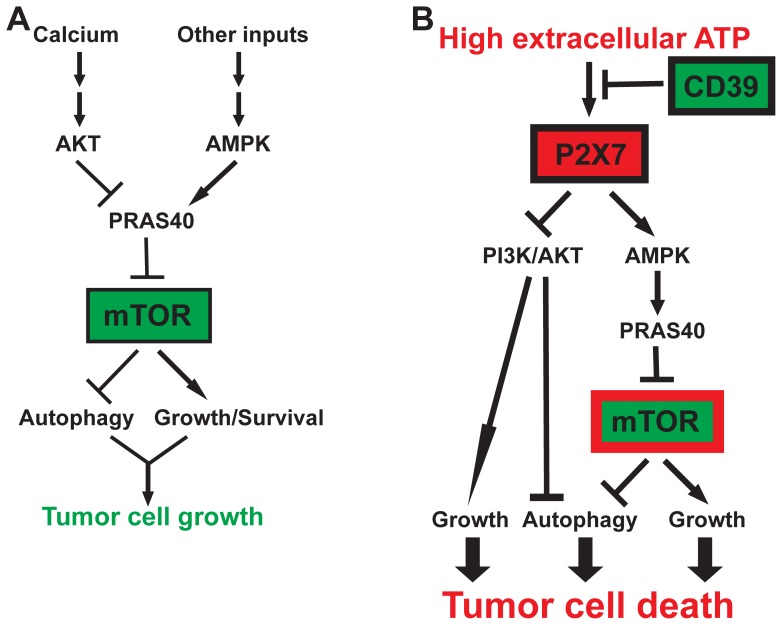
References
-
- White N, Burnstock G (2006) P2 receptors and cancer. Trends Pharmacol Sci 27: 211–217. - PubMed
-
- Eltzschig H, Sitkovsky M, Robson SC (2012) Purinergic signaling during Inflammation. N Engl J Med In press. - PubMed
-
- Volonte C, Amadio S, D’Ambrosi N, Colpi M, Burnstock G (2006) P2 receptor web: complexity and fine-tuning. Pharmacol Ther 112: 264–280. - PubMed
-
- Khakh BS, North RA (2006) P2X receptors as cell-surface ATP sensors in health and disease. Nature 442: 527–532. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
