

Gap junctions and Bystander Effects: Good Samaritans and executioners
- PMID: 23565352
- PMCID: PMC3614363
- DOI: 10.1002/wmts.72
Gap junctions and Bystander Effects: Good Samaritans and executioners
Abstract
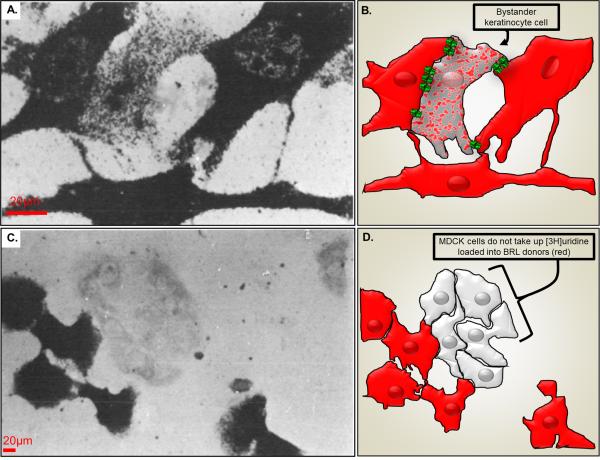
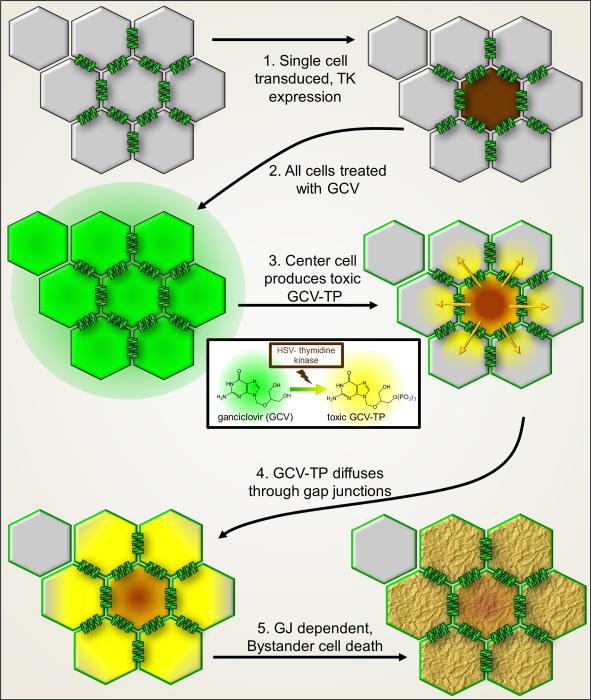
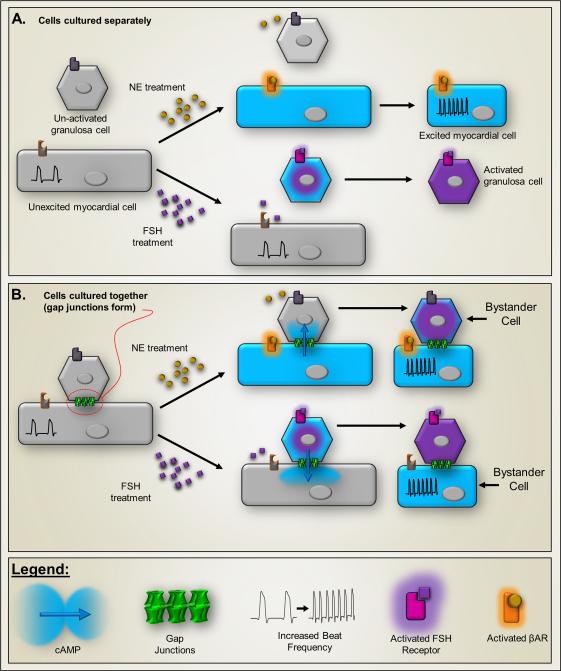
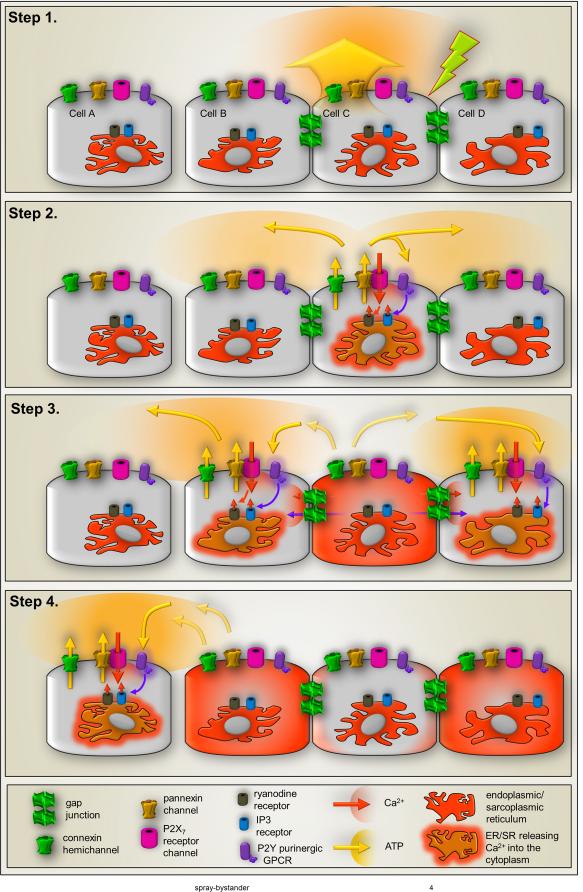
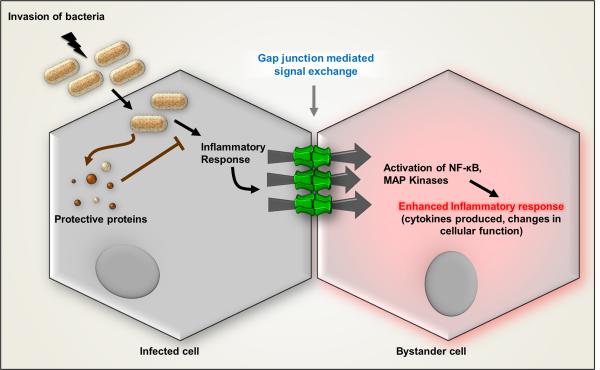
The "Bystander" and "Good Samaritan" effects involve the transfer of toxic or beneficial compounds from one cell to a generally adjacent other through gap junction channels and through extracellular routes. The variety of injuries in which bystander cell killing or protection occurs has greatly expanded in the last decade to include infectious agents and therapeutic compounds, radiation injury, chaperones in cell therapy and apoptosis in development. This has been accompanied by the appreciation that both gap junction mediated and paracrine routes are used for the signaling of the "kiss of life" and the "kiss of death" and that manipulations of these pathways and the molecules that use them may find therapeutic utility in treatment of a variety of pathological conditions.
Figures
References
-
- Pitts JD. The discovery of metabolic co-operation. Bioessays. 1998;20:1047–51. - PubMed
-
- Kimura S, Hase K, Ohno H. Tunneling nanotubes: emerging view of their molecular components and formation mechanisms. Experimental cell research. 2012;318:1699–706. - PubMed
-
- Hamada N, Maeda M, Otsuka K, Tomita M. Signaling pathways underpinning the manifestations of ionizing radiation-induced bystander effects. Curr Mol Pharmacol. 2011;4:79–95. - PubMed
-
- Yang L, Cranson D, Trinkaus-Randall V. Cellular injury induces activation of MAPK via P2Y receptors. J Cell Biochem. 2004;91:938–50. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous