

Targeting of the small GTPase Rab6A' by the Legionella pneumophila effector LidA
- PMID: 23569112
- PMCID: PMC3676037
- DOI: 10.1128/IAI.00157-13
Targeting of the small GTPase Rab6A' by the Legionella pneumophila effector LidA
Abstract
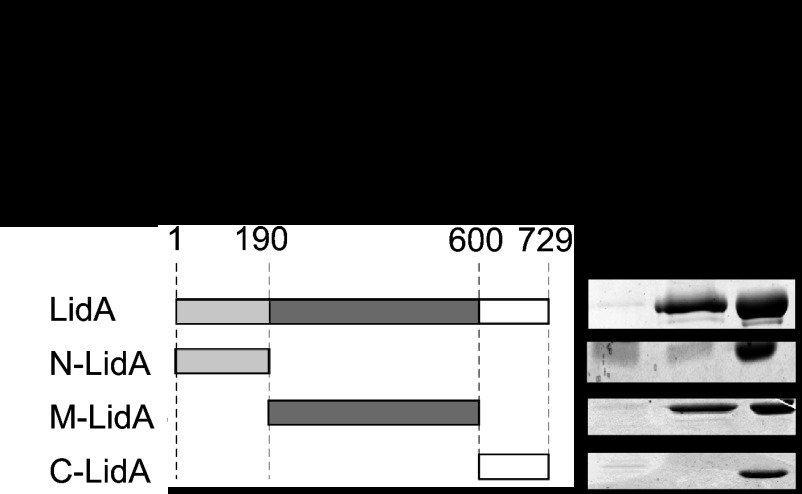
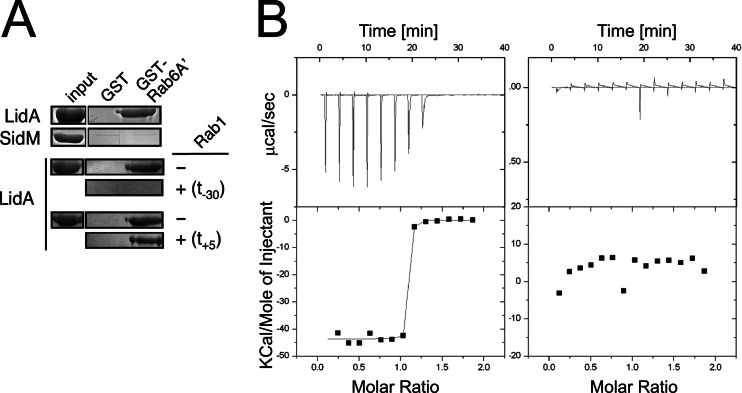
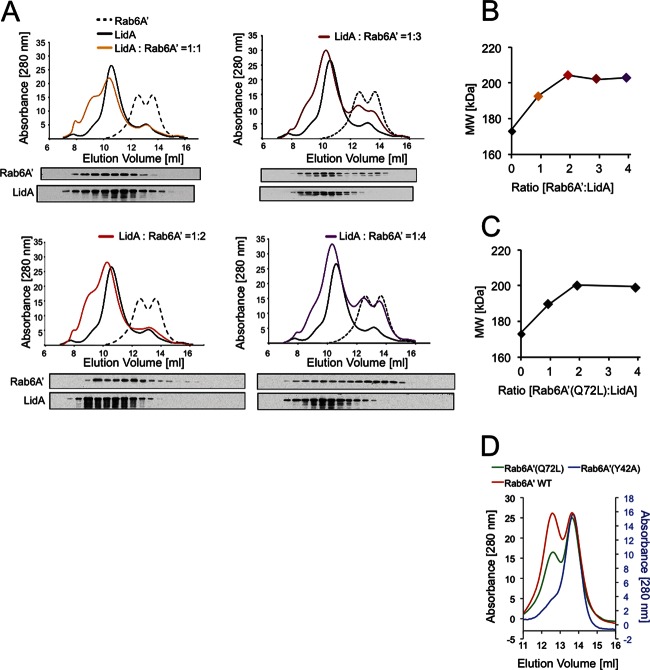
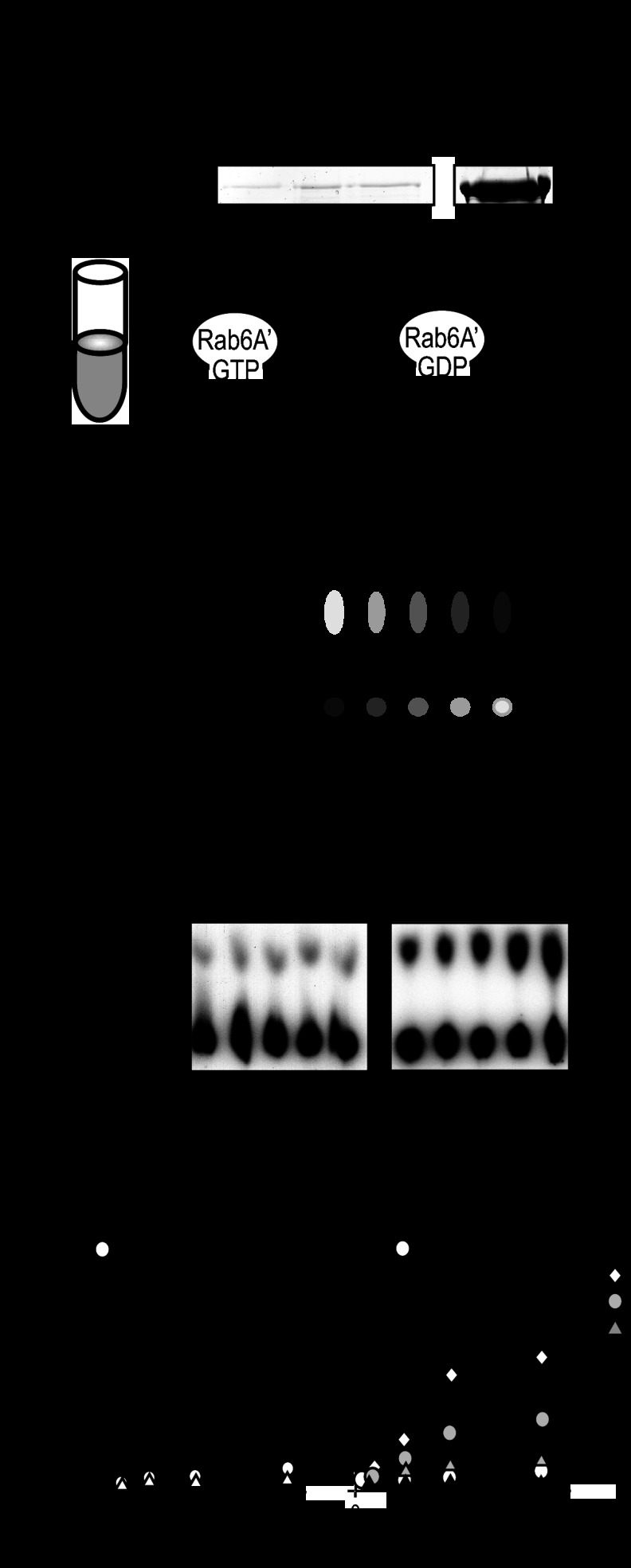
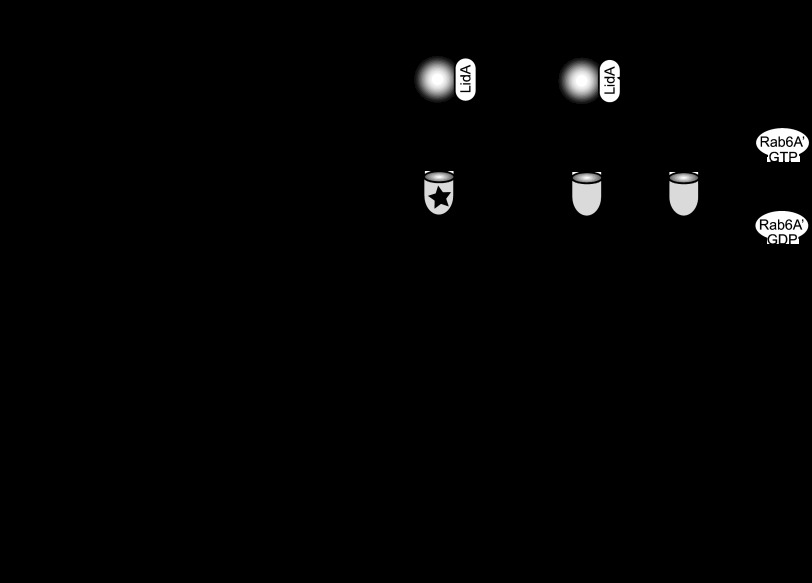
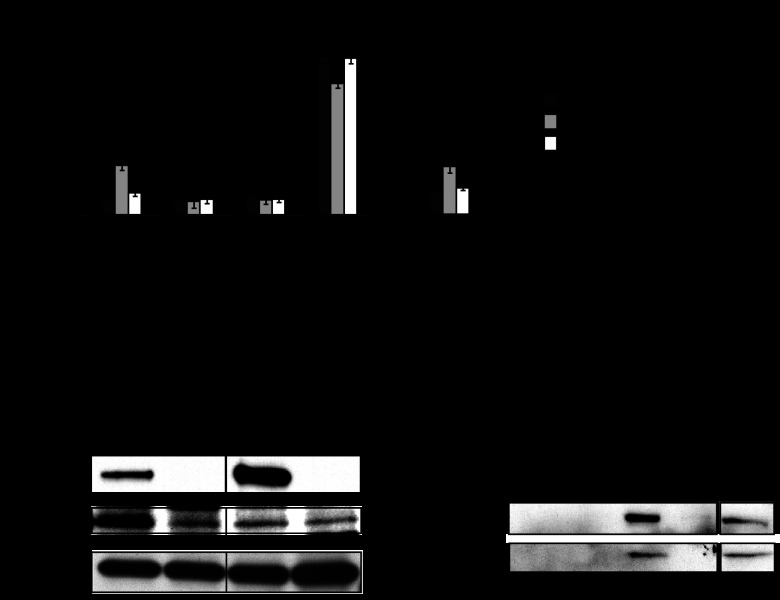
When the bacterium Legionella pneumophila, the causative agent of Legionnaires' disease, is phagocytosed by alveolar macrophages, it delivers a large number of effector proteins through its Dot/Icm type IV secretion system into the host cell cytosol. Among those proteins is LidA, an effector that interacts with several host GTPases of the Rab family, including Rab6A', a regulator of retrograde vesicle trafficking within eukaryotic cells. The effect of LidA on Rab6A' function and the role of Rab6A' for L. pneumophila growth within host cells has been unclear. Here, we show that LidA preferentially binds Rab6A' in the active GTP-bound conformation. Rab6 binding occurred through the central region of LidA and followed a stoichiometry for LidA and Rab6A' of 1:2. LidA maintained Rab6A' in the active conformation by efficiently blocking the hydrolysis of GTP by Rab6A', even in the presence of cellular GTPase-activating proteins, suggesting that the function of Rab6A' must be important for efficient intracellular replication of L. pneumophila. Accordingly, we found that production of constitutively inactive Rab6A'(T27N) but not constitutively active Rab6A'(Q72L) significantly reduced the ability of L. pneumophila to initiate intracellular replication in human macrophages. Thus, the presence of an active pool of Rab6 within host cells early during infection is required to support efficient intracellular growth of L. pneumophila.
Figures
References
-
- Fraser DW, Tsai TR, Orenstein W, Parkin WE, Beecham HJ, Sharrar RG, Harris J, Mallison GF, Martin SM, McDade JE, Shepard CC, Brachman PS. 1977. Legionnaires' disease: description of an epidemic of pneumonia. N. Engl. J. Med. 297:1189–1197 - PubMed
-
- McDade JE, Shepard CC, Fraser DW, Tsai TR, Redus MA, Dowdle WR. 1977. Legionnaires' disease: isolation of a bacterium and demonstration of its role in other respiratory disease. N. Engl. J. Med. 297:1197–1203 - PubMed
-
- Segal G, Shuman HA. 1998. Intracellular multiplication and human macrophage killing by Legionella pneumophila are inhibited by conjugal components of IncQ plasmid RSF1010. Mol. Microbiol. 30:197–208 - PubMed
-
- Vogel JP, Andrews HL, Wong SK, Isberg RR. 1998. Conjugative transfer by the virulence system of Legionella pneumophila. Science 279:873–876 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
