

Molecular mechanisms for the subversion of MyD88 signaling by TcpC from virulent uropathogenic Escherichia coli
- PMID: 23569230
- PMCID: PMC3637702
- DOI: 10.1073/pnas.1215770110
Molecular mechanisms for the subversion of MyD88 signaling by TcpC from virulent uropathogenic Escherichia coli
Abstract
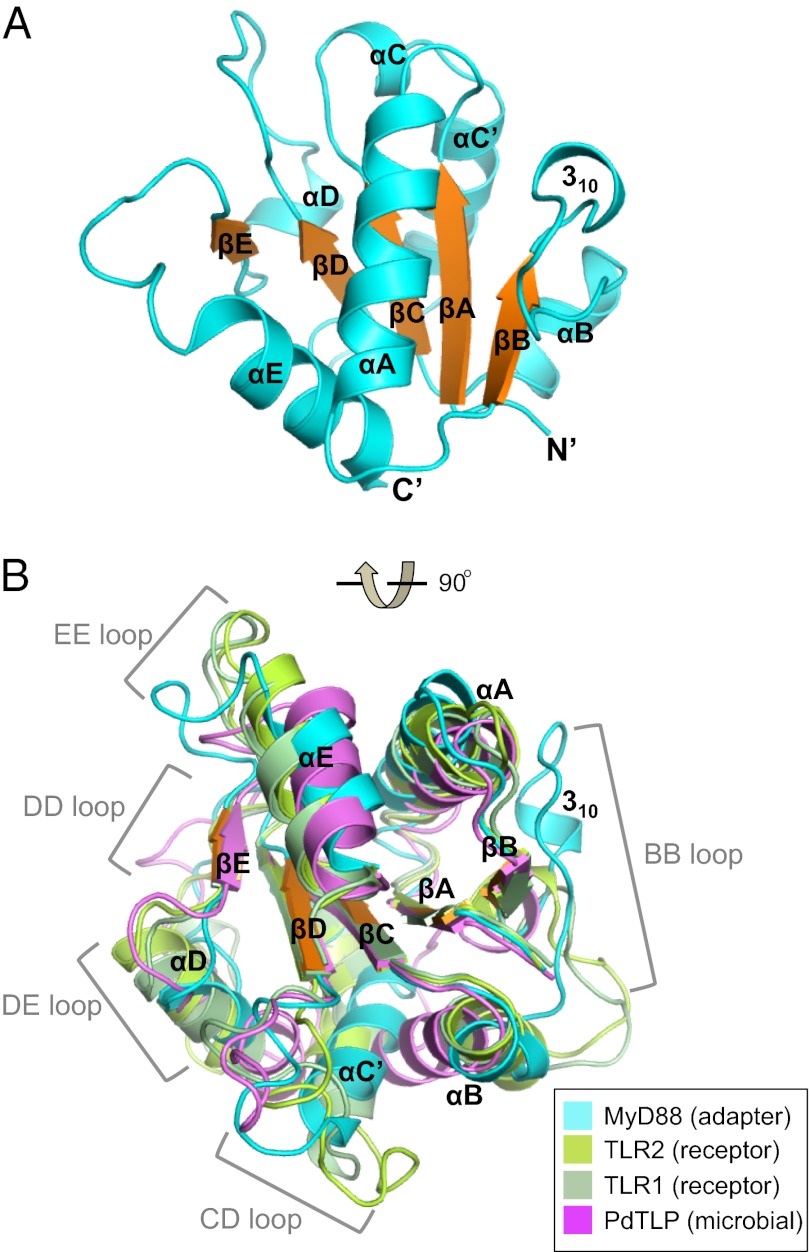
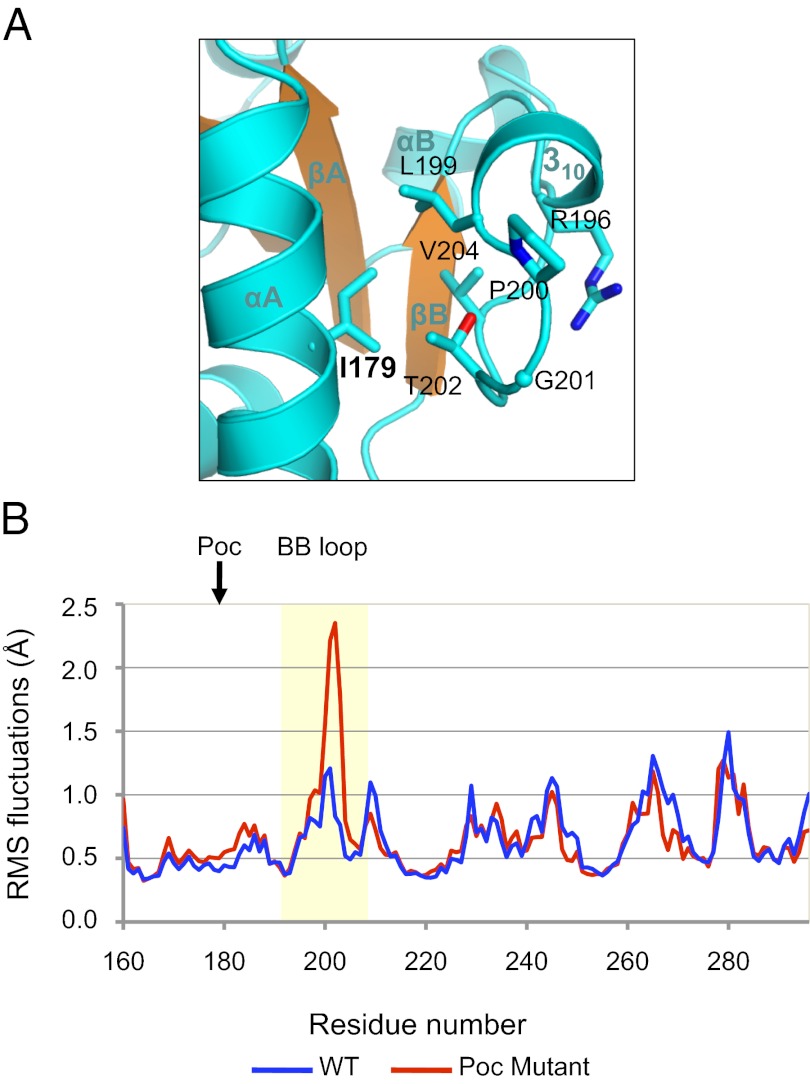
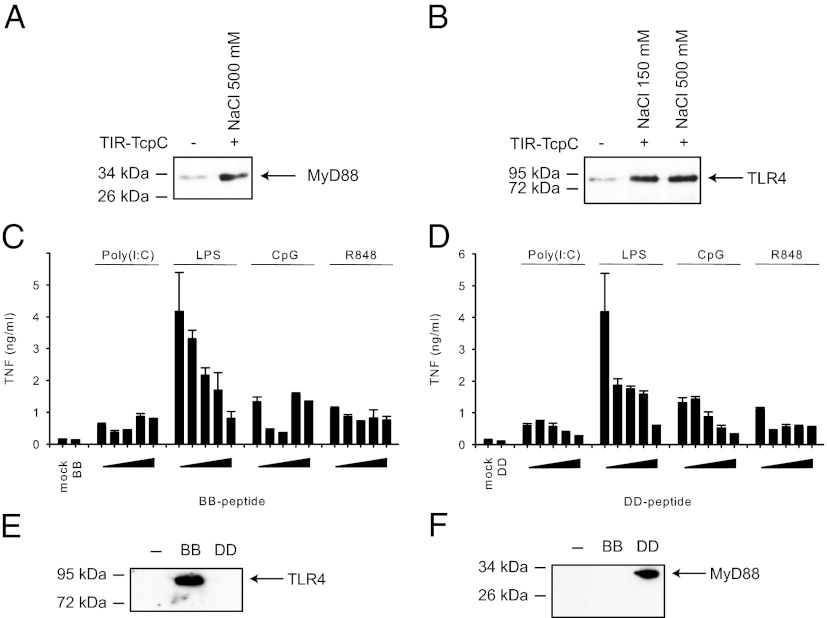
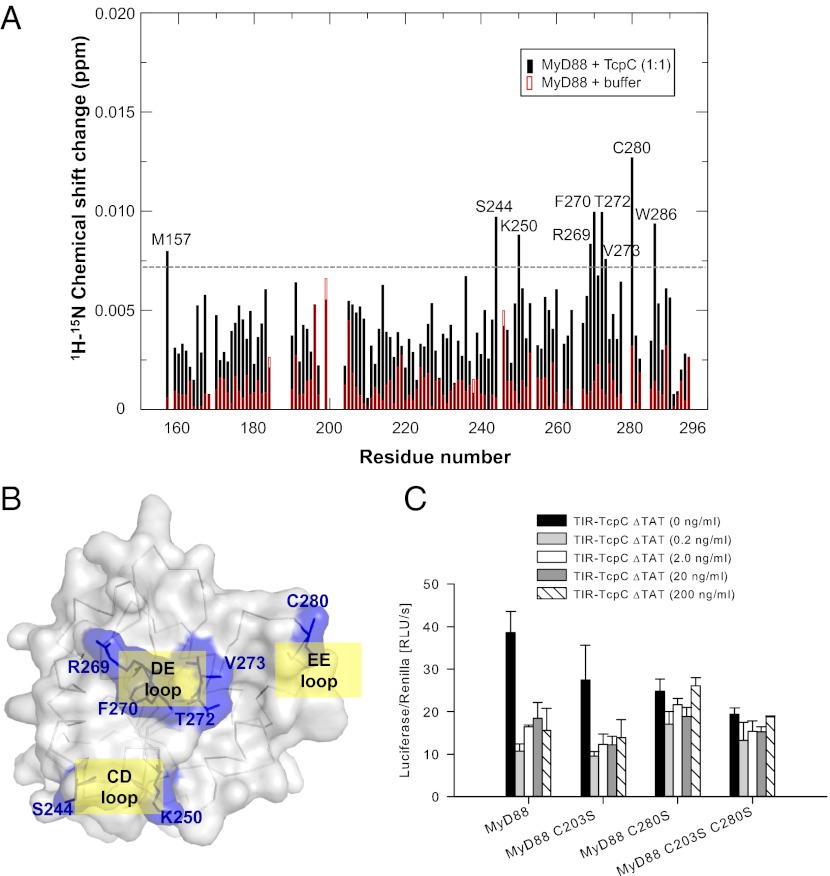
The Toll/IL-1 receptor (TIR) domains are crucial signaling modules during innate immune responses involving the Toll-like receptors (TLRs) and IL-1 receptor (IL-1R). Myeloid differential factor 88 (MyD88) is a central TIR domain-containing adapter molecule responsible for nearly all TLR-mediated signaling and is targeted by a TIR domain-containing protein C (TcpC) from virulent uropathogenic Escherichia coli, a common human pathogen. The mechanism of such molecular antagonism has remained elusive. We present the crystal structure of the MyD88 TIR domain with distinct loop conformations that underscore the functional specialization of the adapter, receptor, and microbial TIR domains. Our structural analyses shed light on the genetic mutations at these loops as well as the Poc site. We demonstrate that TcpC directly associates with MyD88 and TLR4 through its predicted DD and BB loops to impair the TLR-induced cytokine induction. Furthermore, NMR titration experiments identify the unique CD, DE, and EE loops from MyD88 at the TcpC-interacting surface, suggesting that TcpC specifically engages these MyD88 structural elements for immune suppression. These findings thus provide a molecular basis for the subversion of TLR signaling by the uropathogenic E. coli virulence factor TcpC and furnish a framework for the design of novel therapeutic agents that modulate immune activation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Janeway CA, Jr, Medzhitov R. Innate immune recognition. Annu Rev Immunol. 2002;20:197–216. - PubMed
-
- Akira S, Takeda K. Toll-like receptor signalling. Nat Rev Immunol. 2004;4(7):499–511. - PubMed
-
- O’Neill LAJ, Bowie AG. The family of five: TIR-domain-containing adaptors in Toll-like receptor signalling. Nat Rev Immunol. 2007;7(5):353–364. - PubMed
-
- Kawai T, Adachi O, Ogawa T, Takeda K, Akira S. Unresponsiveness of MyD88-deficient mice to endotoxin. Immunity. 1999;11(1):115–122. - PubMed
-
- Adachi O, et al. Targeted disruption of the MyD88 gene results in loss of IL-1- and IL-18-mediated function. Immunity. 1998;9(1):143–150. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
