

Coordinated transcriptional regulation of bone homeostasis by Ebf1 and Zfp521 in both mesenchymal and hematopoietic lineages
- PMID: 23569325
- PMCID: PMC3646489
- DOI: 10.1084/jem.20121187
Coordinated transcriptional regulation of bone homeostasis by Ebf1 and Zfp521 in both mesenchymal and hematopoietic lineages
Abstract
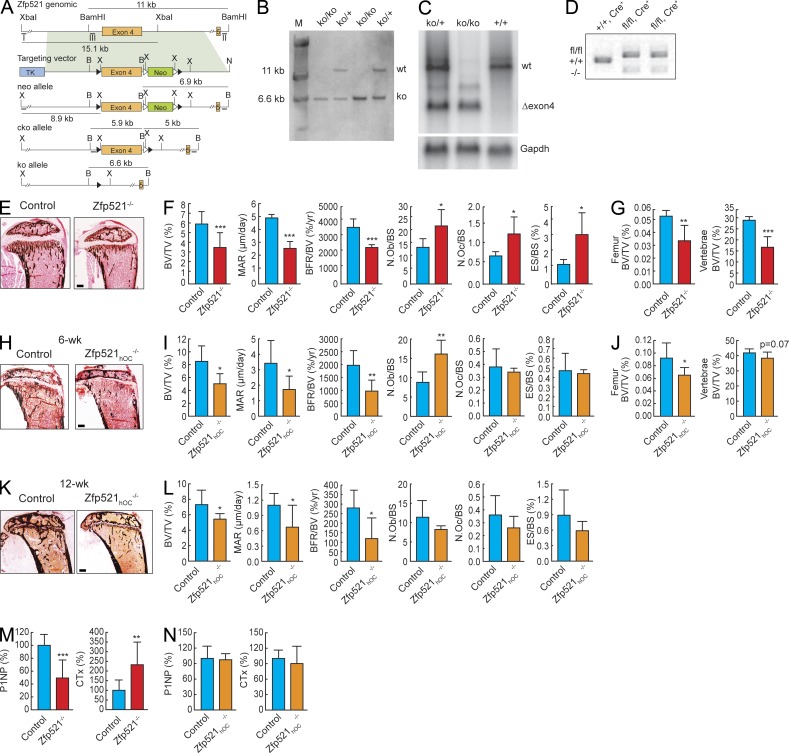
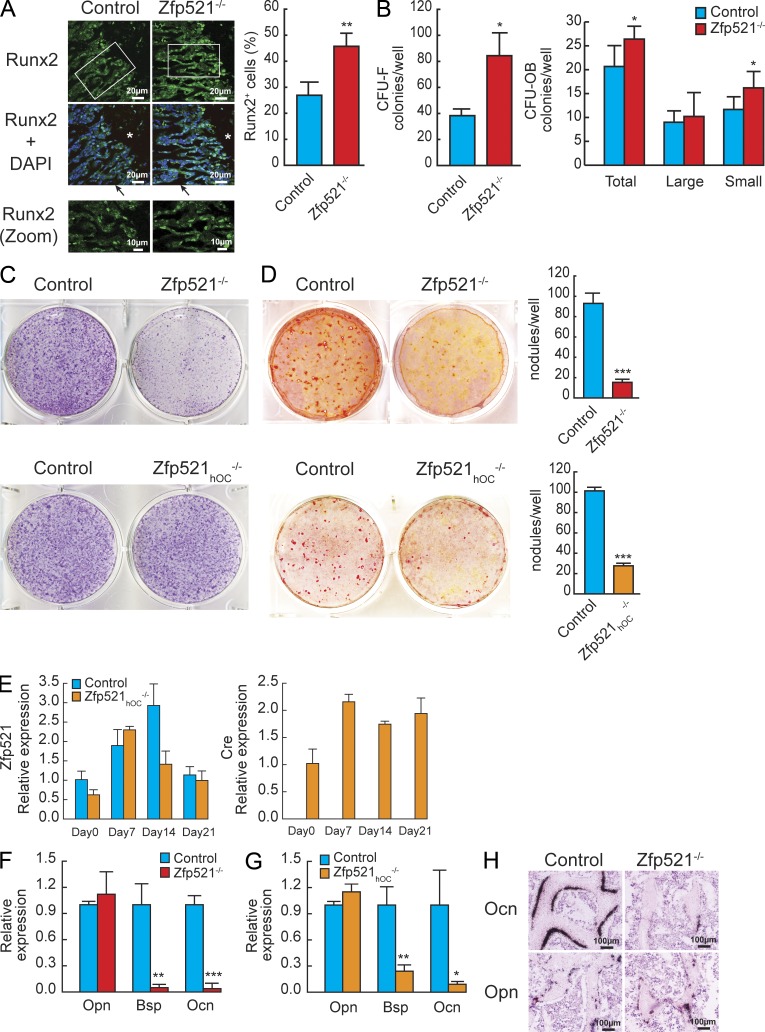
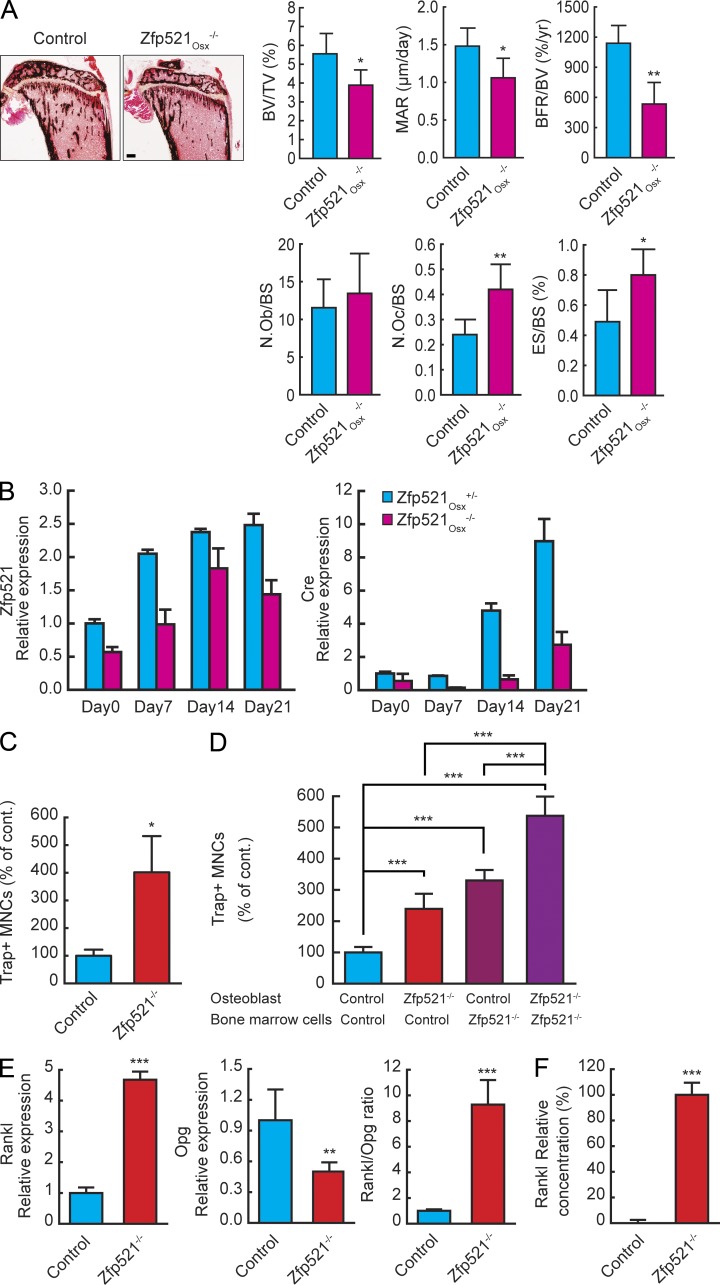
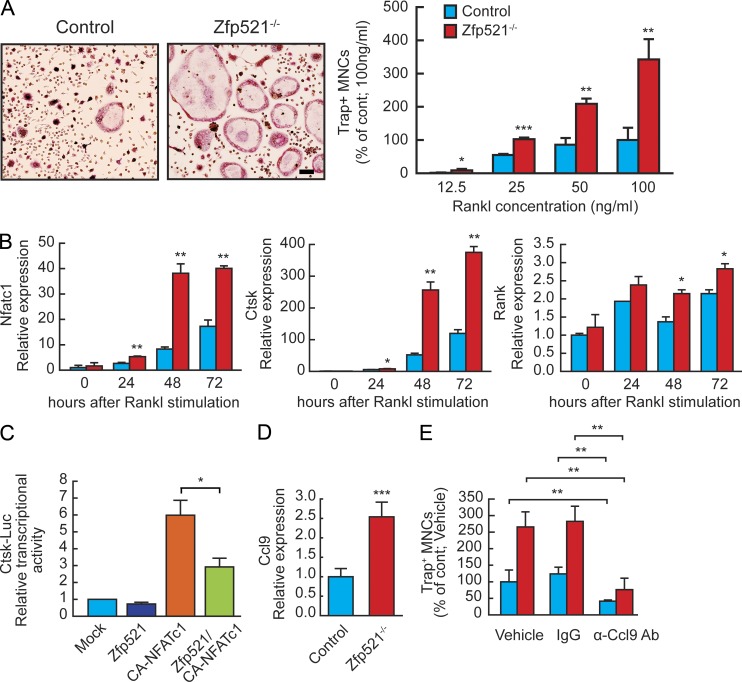
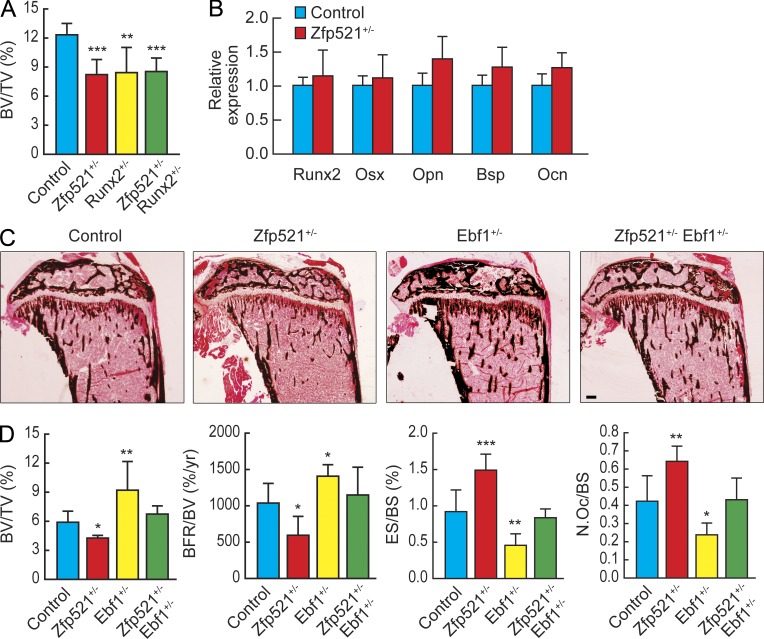
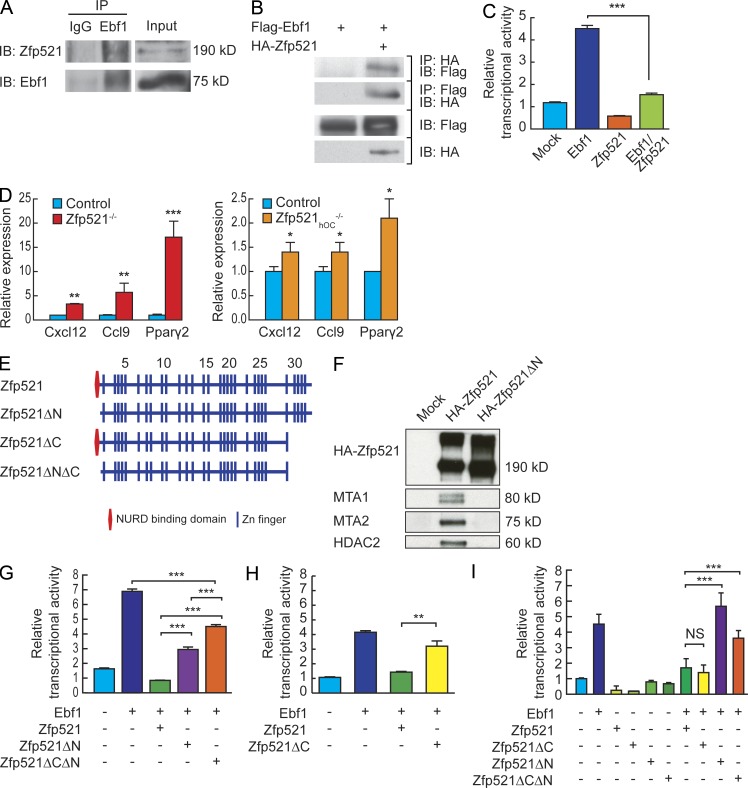
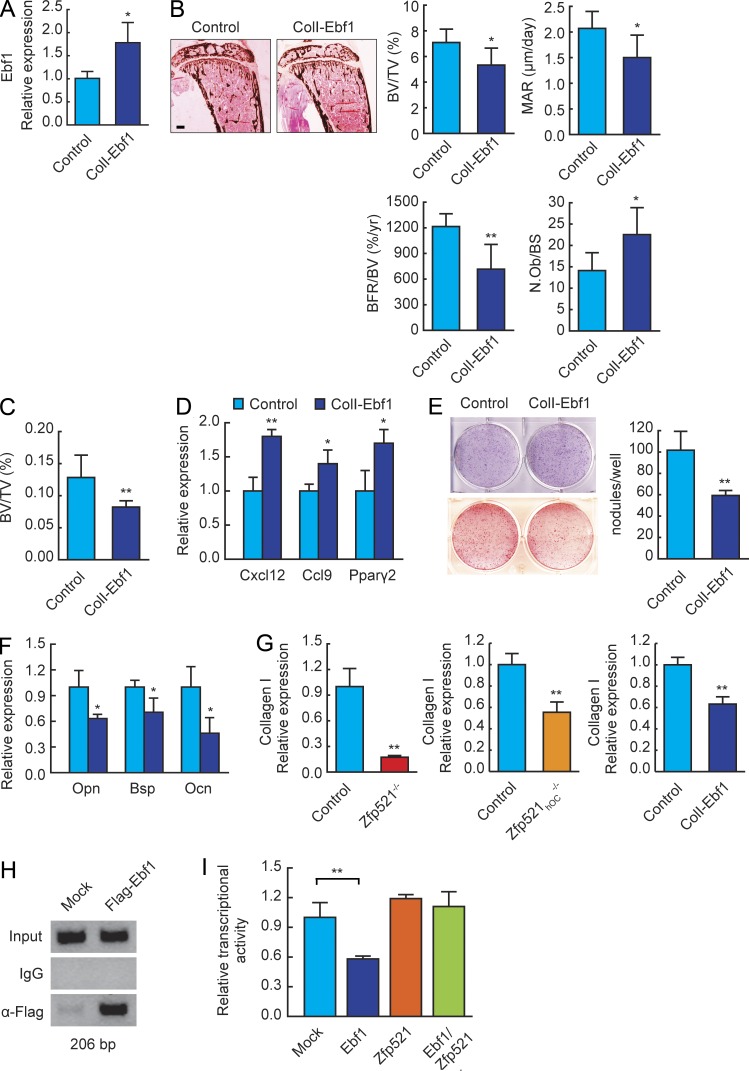
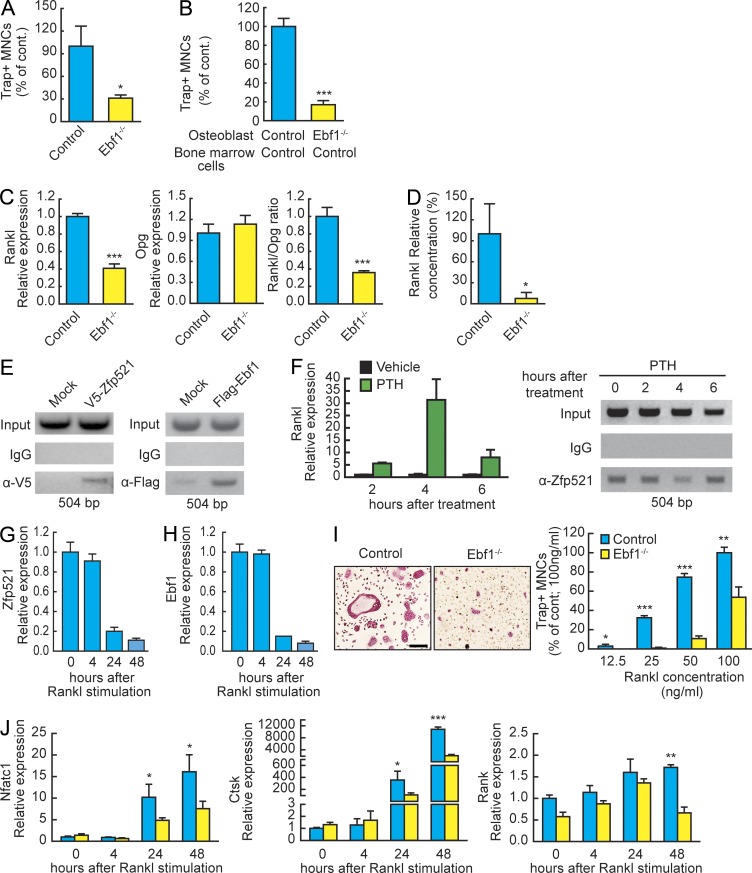
Bone homeostasis is maintained by the coupled actions of hematopoietic bone-resorbing osteoclasts (OCs) and mesenchymal bone-forming osteoblasts (OBs). Here we identify early B cell factor 1 (Ebf1) and the transcriptional coregulator Zfp521 as components of the machinery that regulates bone homeostasis through coordinated effects in both lineages. Deletion of Zfp521 in OBs led to impaired bone formation and increased OB-dependent osteoclastogenesis (OC-genesis), and deletion in hematopoietic cells revealed a strong cell-autonomous role for Zfp521 in OC progenitors. In adult mice, the effects of Zfp521 were largely caused by repression of Ebf1, and the bone phenotype of Zfp521(+/-) mice was rescued in Zfp521(+/-):Ebf1(+/-) mice. Zfp521 interacted with Ebf1 and repressed its transcriptional activity. Accordingly, deletion of Zfp521 led to increased Ebf1 activity in OBs and OCs. In vivo, Ebf1 overexpression in OBs resulted in suppressed bone formation, similar to the phenotype seen after OB-targeted deletion of Zfp521. Conversely, Ebf1 deletion led to cell-autonomous defects in both OB-dependent and cell-intrinsic OC-genesis, a phenotype opposite to that of the Zfp521 knockout. Thus, we have identified the interplay between Zfp521 and Ebf1 as a novel rheostat for bone homeostasis.
Figures
References
-
- Bond H.M., Mesuraca M., Carbone E., Bonelli P., Agosti V., Amodio N., De Rosa G., Di Nicola M., Gianni A.M., Moore M.A., et al. 2004. Early hematopoietic zinc finger protein (EHZF), the human homolog to mouse Evi3, is highly expressed in primitive human hematopoietic cells. Blood. 103:2062–2070 10.1182/blood-2003-07-2388 - DOI - PubMed
-
- Correa D., Hesse E., Seriwatanachai D., Kiviranta R., Saito H., Yamana K., Neff L., Atfi A., Coillard L., Sitara D., et al. 2010. Zfp521 is a target gene and key effector of parathyroid hormone-related peptide signaling in growth plate chondrocytes. Dev. Cell. 19:533–546 10.1016/j.devcel.2010.09.008 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
