

Evolution of homospermidine synthase in the convolvulaceae: a story of gene duplication, gene loss, and periods of various selection pressures
- PMID: 23572540
- PMCID: PMC3663263
- DOI: 10.1105/tpc.113.109744
Evolution of homospermidine synthase in the convolvulaceae: a story of gene duplication, gene loss, and periods of various selection pressures
Abstract
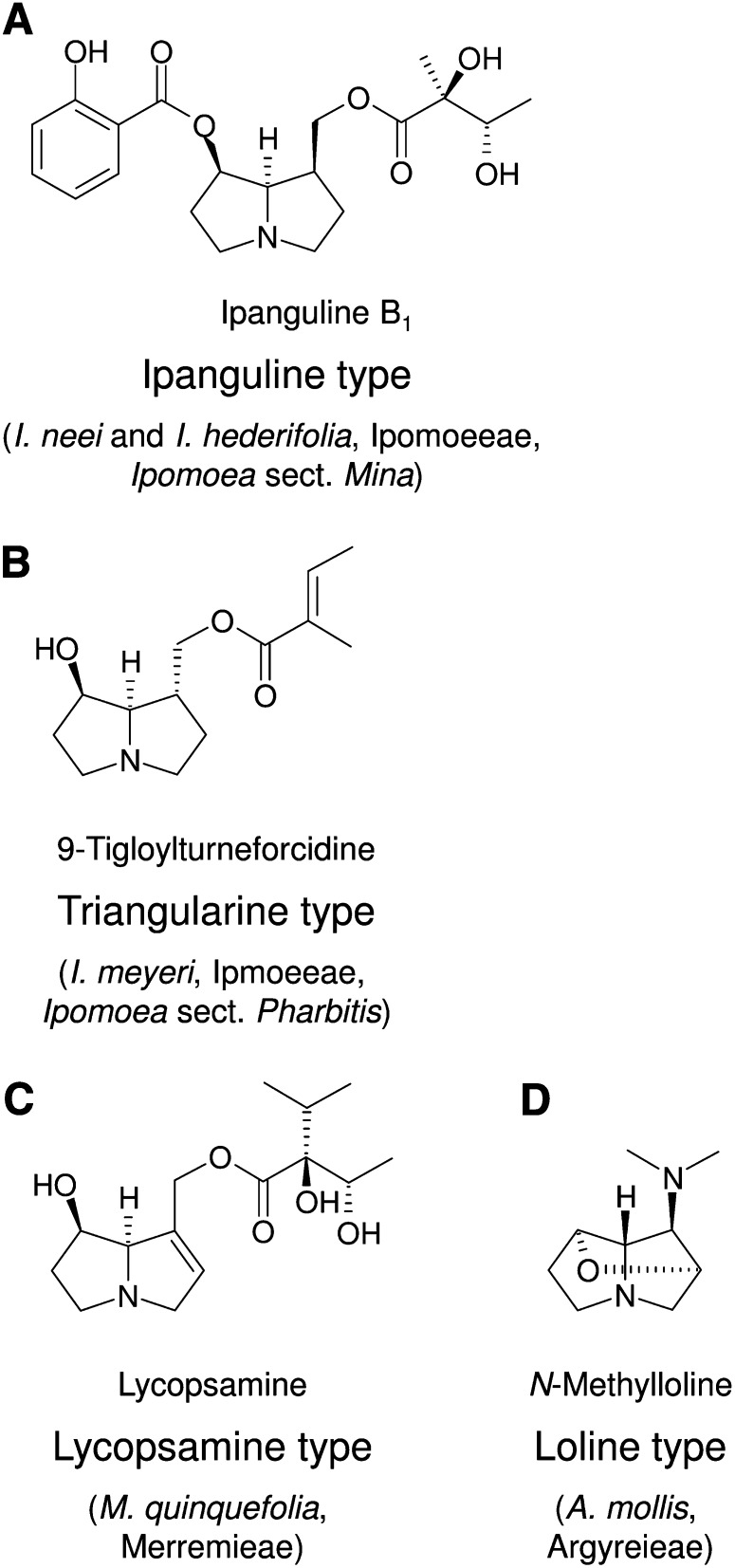
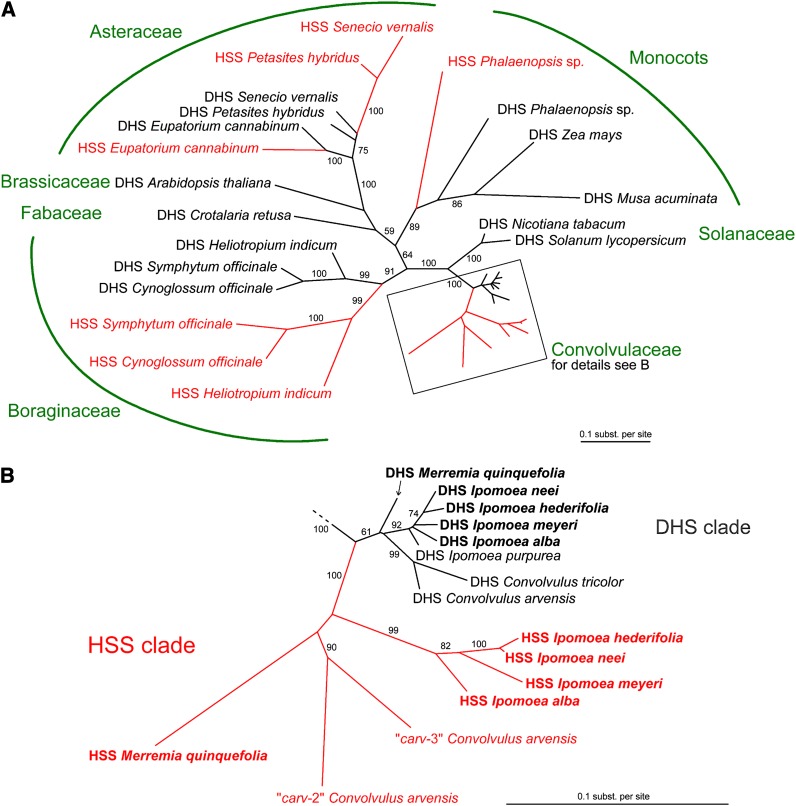
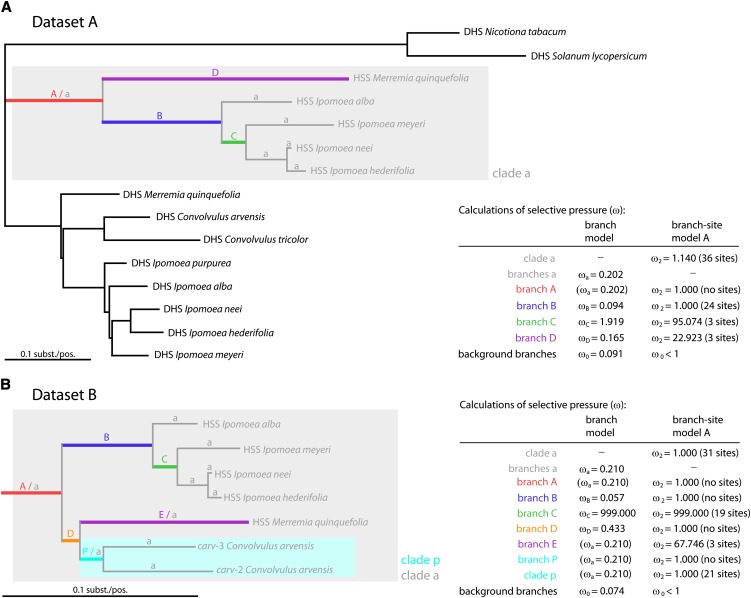
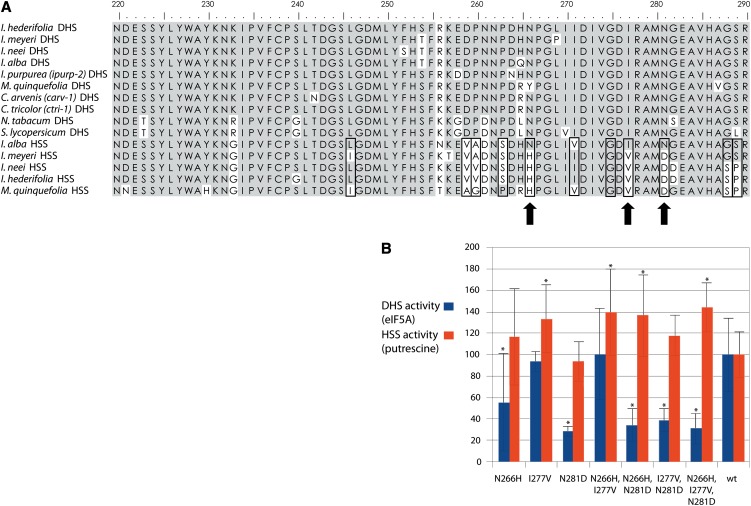
Homospermidine synthase (HSS), the first pathway-specific enzyme of pyrrolizidine alkaloid biosynthesis, is known to have its origin in the duplication of a gene encoding deoxyhypusine synthase. To study the processes that followed this gene duplication event and gave rise to HSS, we identified sequences encoding HSS and deoxyhypusine synthase from various species of the Convolvulaceae. We show that HSS evolved only once in this lineage. This duplication event was followed by several losses of a functional gene copy attributable to gene loss or pseudogenization. Statistical analyses of sequence data suggest that, in those lineages in which the gene copy was successfully recruited as HSS, the gene duplication event was followed by phases of various selection pressures, including purifying selection, relaxed functional constraints, and possibly positive Darwinian selection. Site-specific mutagenesis experiments have confirmed that the substitution of sites predicted to be under positive Darwinian selection is sufficient to convert a deoxyhypusine synthase into a HSS. In addition, analyses of transcript levels have shown that HSS and deoxyhypusine synthase have also diverged with respect to their regulation. The impact of protein-protein interaction on the evolution of HSS is discussed with respect to current models of enzyme evolution.
Figures
References
-
- Anisimova M., Bielawski J.P., Yang Z. (2001). Accuracy and power of the likelihood ratio test in detecting adaptive molecular evolution. Mol. Biol. Evol. 18: 1585–1592 - PubMed
-
- Anisimova M., Yang Z. (2007). Multiple hypothesis testing to detect lineages under positive selection that affects only a few sites. Mol. Biol. Evol. 24: 1219–1228 - PubMed
-
- Barkman T.J. (2003). Evidence for positive selection on the floral scent gene isoeugenol-O-methyltransferase. Mol. Biol. Evol. 20: 168–172 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
