

ApiAP2 transcription factor restricts development of the Toxoplasma tissue cyst
- PMID: 23572590
- PMCID: PMC3637731
- DOI: 10.1073/pnas.1300059110
ApiAP2 transcription factor restricts development of the Toxoplasma tissue cyst
Abstract
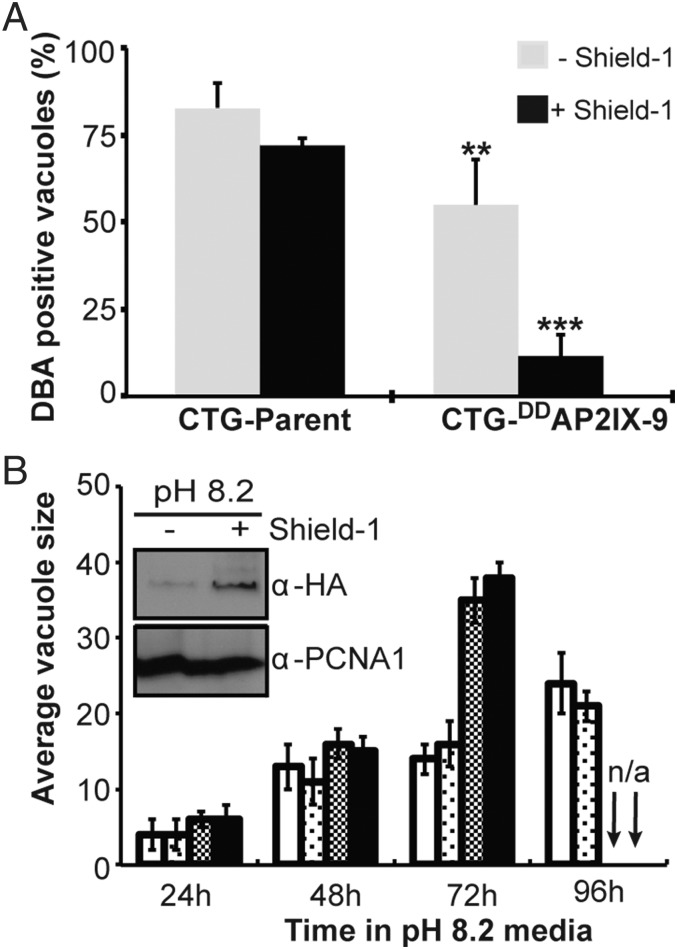
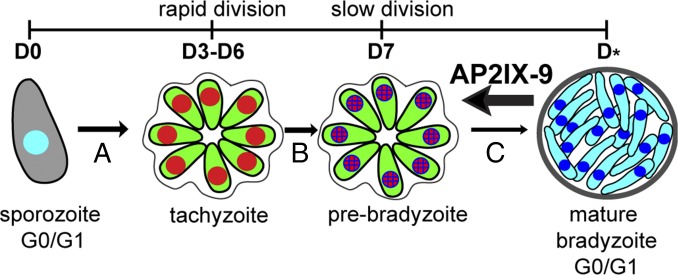
Cellular differentiation leading to formation of the bradyzoite tissue cyst stage is the underlying cause of chronic toxoplasmosis. Consequently, mechanisms responsible for controlling development in the Toxoplasma intermediate life cycle have long been sought. Here, we identified 15 Toxoplasma mRNAs induced in early bradyzoite development that encode proteins with apicomplexan AP2 (ApiAP2) DNA binding domains. Of these 15 mRNAs, the AP2IX-9 mRNA demonstrated the largest expression increase during alkaline-induced differentiation. At the protein level, we found that AP2IX-9 was restricted to the early bradyzoite nucleus and is repressed in tachyzoites and in mature bradyzoites from 30-d infected animals. Conditional overexpression of AP2IX-9 significantly reduced tissue cyst formation and conferred alkaline pH-resistant growth, whereas disruption of the AP2IX-9 gene increased tissue cyst formation, indicating AP2IX-9 operates as a repressor of bradyzoite development. Consistent with a role as a repressor, AP2IX-9 specifically inhibited the expression of bradyzoite mRNAs, including the canonical bradyzoite marker, bradyzoite antigen 1 (BAG1). Using protein binding microarrays, we established the AP2 domain of AP2IX-9 binds a CAGTGT DNA sequence motif and is capable of binding cis-regulatory elements controlling the BAG1 and bradyzoite-specific nucleoside triphosphatase (B-NTPase) promoters. The effect of AP2IX-9 on BAG1 expression was direct because this factor inhibits expression of a firefly luciferase reporter under the control of the BAG1 promoter in vivo, and epitope-tagged AP2IX-9 can be immunoprecipitated with the BAG1 promoter in parasite chromatin. Altogether, these results indicate AP2IX-9 restricts Toxoplasma commitment to develop the mature bradyzoite tissue cyst.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



Similar articles
-
Opposing Transcriptional Mechanisms Regulate Toxoplasma Development.mSphere. 2017 Feb 22;2(1):e00347-16. doi: 10.1128/mSphere.00347-16. eCollection 2017 Jan-Feb. mSphere. 2017. PMID: 28251183 Free PMC article.
-
Toxoplasma gondii AP2IX-4 Regulates Gene Expression during Bradyzoite Development.mSphere. 2017 Mar 15;2(2):e00054-17. doi: 10.1128/mSphere.00054-17. eCollection 2017 Mar-Apr. mSphere. 2017. PMID: 28317026 Free PMC article.
-
The transcription of bradyzoite genes in Toxoplasma gondii is controlled by autonomous promoter elements.Mol Microbiol. 2008 Jun;68(6):1502-18. doi: 10.1111/j.1365-2958.2008.06249.x. Epub 2008 Apr 21. Mol Microbiol. 2008. PMID: 18433450 Free PMC article.
-
The Epigenome, Cell Cycle, and Development in Toxoplasma.Annu Rev Microbiol. 2018 Sep 8;72:479-499. doi: 10.1146/annurev-micro-090817-062741. Epub 2018 Jun 22. Annu Rev Microbiol. 2018. PMID: 29932347 Review.
-
Stress-related and spontaneous stage differentiation of Toxoplasma gondii.Mol Biosyst. 2008 Aug;4(8):824-34. doi: 10.1039/b800520f. Epub 2008 Jun 2. Mol Biosyst. 2008. PMID: 18633484 Review.
Cited by
-
Not your Mother's MAPKs: Apicomplexan MAPK function in daughter cell budding.PLoS Pathog. 2022 Oct 13;18(10):e1010849. doi: 10.1371/journal.ppat.1010849. eCollection 2022 Oct. PLoS Pathog. 2022. PMID: 36227859 Free PMC article. Review.
-
A novel GCN5b lysine acetyltransferase complex associates with distinct transcription factors in the protozoan parasite Toxoplasma gondii.Mol Biochem Parasitol. 2019 Sep;232:111203. doi: 10.1016/j.molbiopara.2019.111203. Epub 2019 Aug 2. Mol Biochem Parasitol. 2019. PMID: 31381949 Free PMC article.
-
ApiAP2 Transcription Factors in Apicomplexan Parasites.Pathogens. 2019 Apr 7;8(2):47. doi: 10.3390/pathogens8020047. Pathogens. 2019. PMID: 30959972 Free PMC article. Review.
-
Efficient Single-Gene and Gene Family Editing in the Apicomplexan Parasite Eimeria tenella Using CRISPR-Cas9.Front Bioeng Biotechnol. 2020 Feb 25;8:128. doi: 10.3389/fbioe.2020.00128. eCollection 2020. Front Bioeng Biotechnol. 2020. PMID: 32158750 Free PMC article.
-
The Cryptosporidium parvum ApiAP2 gene family: insights into the evolution of apicomplexan AP2 regulatory systems.Nucleic Acids Res. 2014 Jul;42(13):8271-84. doi: 10.1093/nar/gku500. Epub 2014 Jun 23. Nucleic Acids Res. 2014. PMID: 24957599 Free PMC article.
References
-
- Dubey JP, Miller NL, Frenkel JK. Toxoplasma gondii life cycle in cats. J Am Vet Med Assoc. 1970;157(11):1767–1770. - PubMed
-
- Kruszon-Moran D, McQuillan GM. Seroprevalence of six infectious diseases among adults in the United States by race/ethnicity: Data from the third national health and nutrition examination survey, 1988--94. Adv Data. 2005;(352):1–9. - PubMed
-
- Jones JL, Kruszon-Moran D, Sanders-Lewis K, Wilson M. Toxoplasma gondii infection in the United States, 1999 2004, decline from the prior decade. Am J Trop Med Hyg. 2007;77(3):405–410. - PubMed
-
- Adomako-Ankomah Y, Wier GM, Boyle JP. Beyond the genome: Recent advances in Toxoplasma gondii functional genomics. Parasite Immunol. 2012;34(2-3):80–89. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
