

Histopathologies, immunolocalization, and a glycan binding screen provide insights into Plasmodium falciparum interactions with the human placenta
- PMID: 23575149
- PMCID: PMC4070867
- DOI: 10.1095/biolreprod.112.106195
Histopathologies, immunolocalization, and a glycan binding screen provide insights into Plasmodium falciparum interactions with the human placenta
Abstract
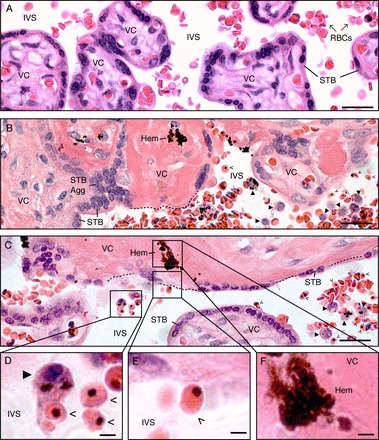
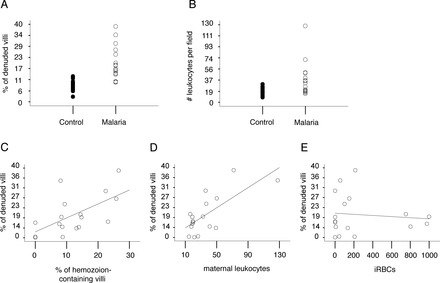
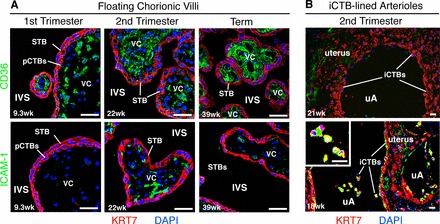
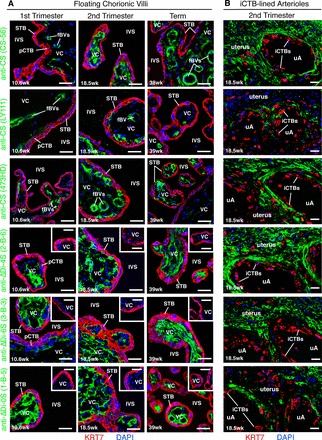
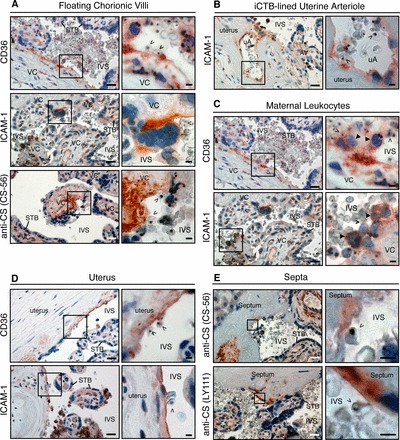
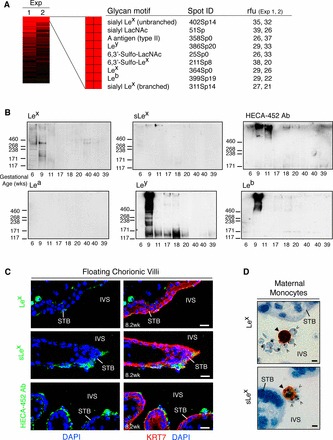
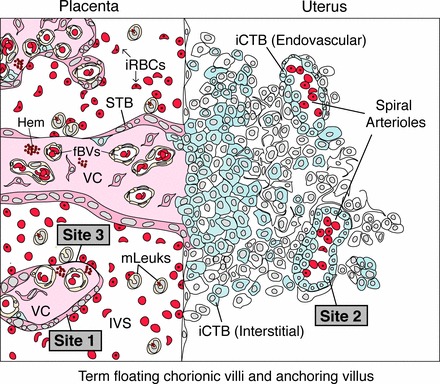
During pregnancy, Plasmodium falciparum-infected erythrocytes cytoadhere to the placenta. Infection is likely initiated at two sites where placental trophoblasts contact maternal blood: 1) via syncytiotrophoblast (STB), a multicellular transporting and biosynthetic layer that forms the surface of chorionic villi and lines the intervillous space, and 2) through invasive cytotrophoblasts, which line uterine vessels that divert blood to the placenta. Here, we investigated mechanisms of infected erythrocyte sequestration in relationship to the microanatomy of the maternal-fetal interface. Histological analyses revealed STB denudation in placental malaria, which brought the stromal cores of villi in direct contact with maternal blood. STB denudation was associated with hemozoin deposition (P = 0.01) and leukocyte infiltration (P = 0.001) and appeared to be a feature of chronic placental malaria. Immunolocalization of infected red blood cell receptors (CD36, ICAM1/CD54, and chondroitin sulfate A) in placentas from uncomplicated pregnancies showed that STB did not stain, while the underlying villous stroma was immunopositive. Invasive cytotrophoblasts expressed ICAM1. In malaria, STB denudation exposed CD36 and chondroitin sulfate A in the villous cores to maternal blood, and STB expressed ICAM1. Finally, we investigated infected erythrocyte adherence to novel receptors by screening an array of 377 glycans. Infected erythrocytes bound Lewis antigens that immunolocalized to STB. Our results suggest that P. falciparum interactions with STB-associated Lewis antigens could initiate placental malaria. Subsequent pathologies, which expose CD36, ICAM1, and chondroitin sulfate A, might propagate the infection.
Keywords: placenta; pregnancy; stroma; syncytiotrophoblast; trophoblast.
Figures
References
-
- Desai M, ter Kuile FO, Nosten F, McGready R, Asamoa K, Brabin B, Newman RD. Epidemiology and burden of malaria in pregnancy. Lancet Infect Dis 2007; 7: 93 104. - PubMed
-
- Rogerson SJ, Mwapasa V, Meshnick SR. Malaria in pregnancy: linking immunity and pathogenesis to prevention. Am J Trop Med Hyg 2007; 77: 14 22. - PubMed
-
- Rogerson SJ, Hviid L, Duffy PE, Leke RF, Taylor DW. Malaria in pregnancy: pathogenesis and immunity. Lancet Infect Dis 2007; 7: 105 117. - PubMed
-
- Brabin BJ, Romagosa C, Abdelgalil S, Menendez C, Verhoeff FH, McGready R, Fletcher KA, Owens S, D'Alessandro U, Nosten F, Fischer PR, Ordi J. The sick placenta—the role of malaria. Placenta 2004; 25: 359 378. - PubMed
-
- Brabin B, Johnson P. Placental malaria and pre-eclampsia through the looking glass backwards? J Reprod Immunol 2005; 65: 1 15. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
