

Overexpression of full-length centrobin rescues limb malformation but not male fertility of the hypodactylous (hd) rats
- PMID: 23577170
- PMCID: PMC3620055
- DOI: 10.1371/journal.pone.0060859
Overexpression of full-length centrobin rescues limb malformation but not male fertility of the hypodactylous (hd) rats
Abstract
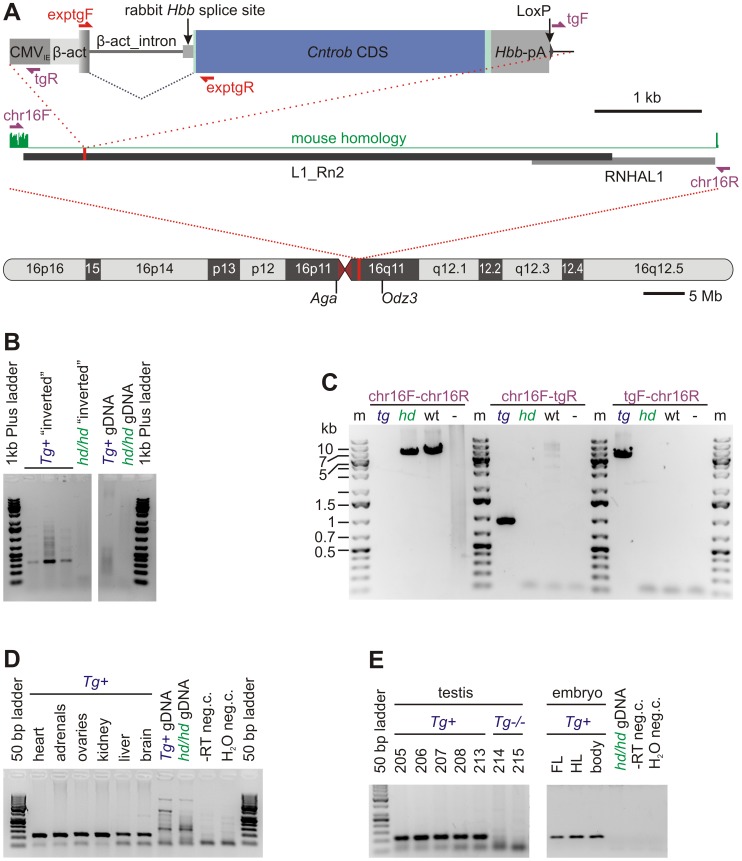
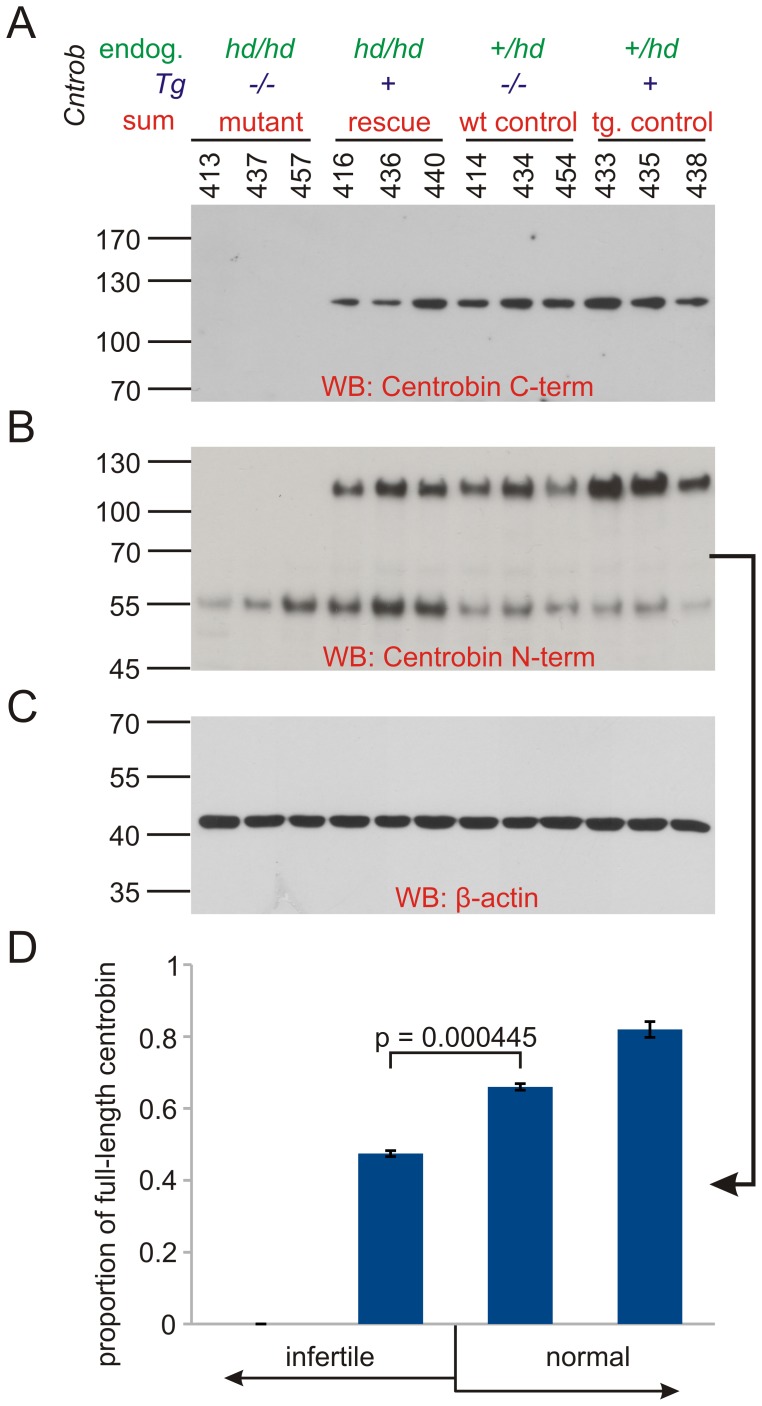
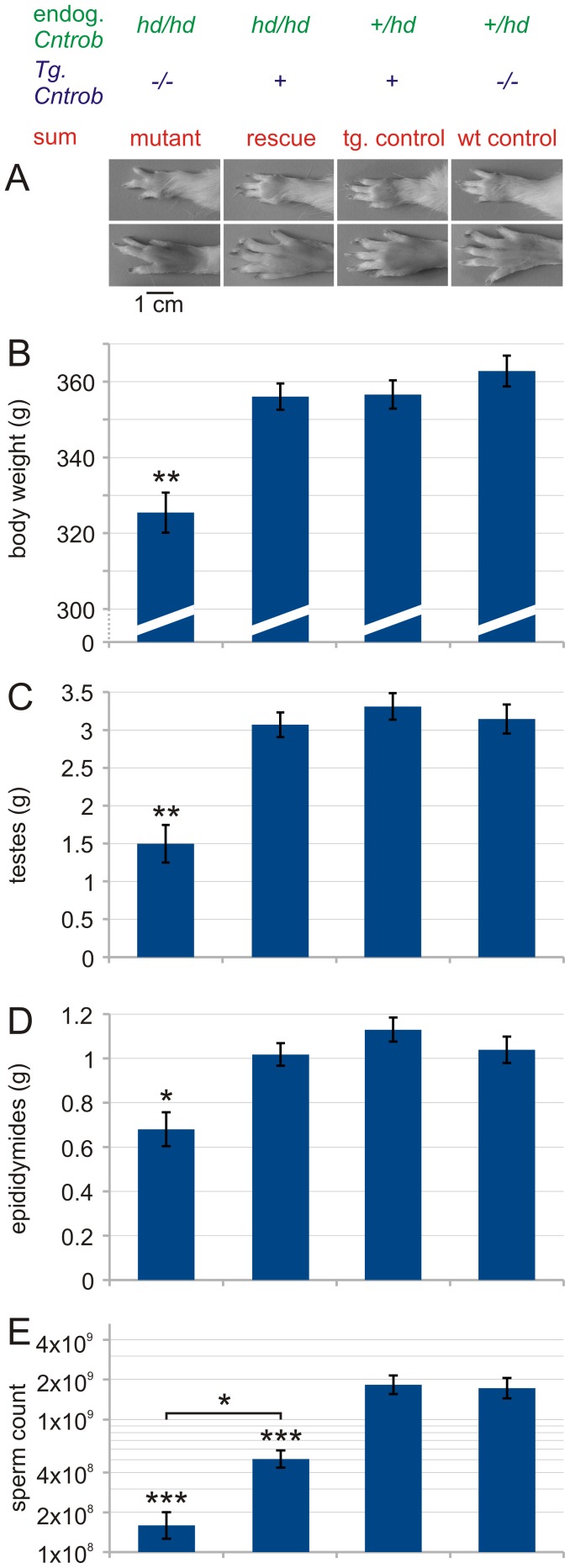
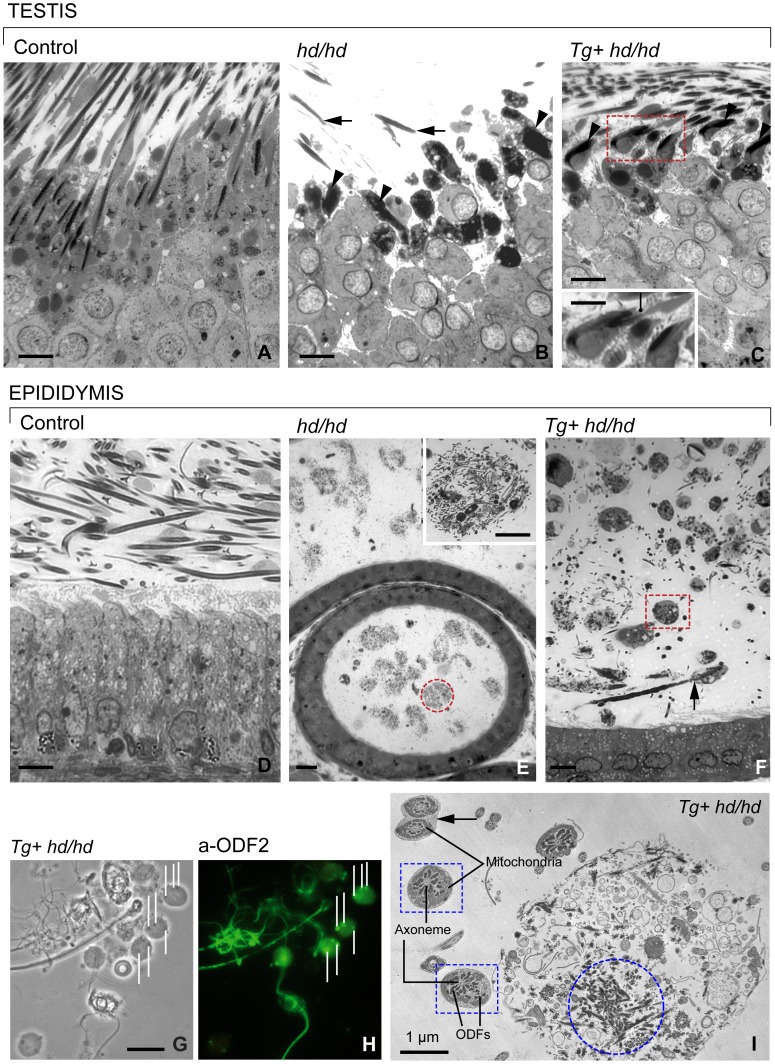
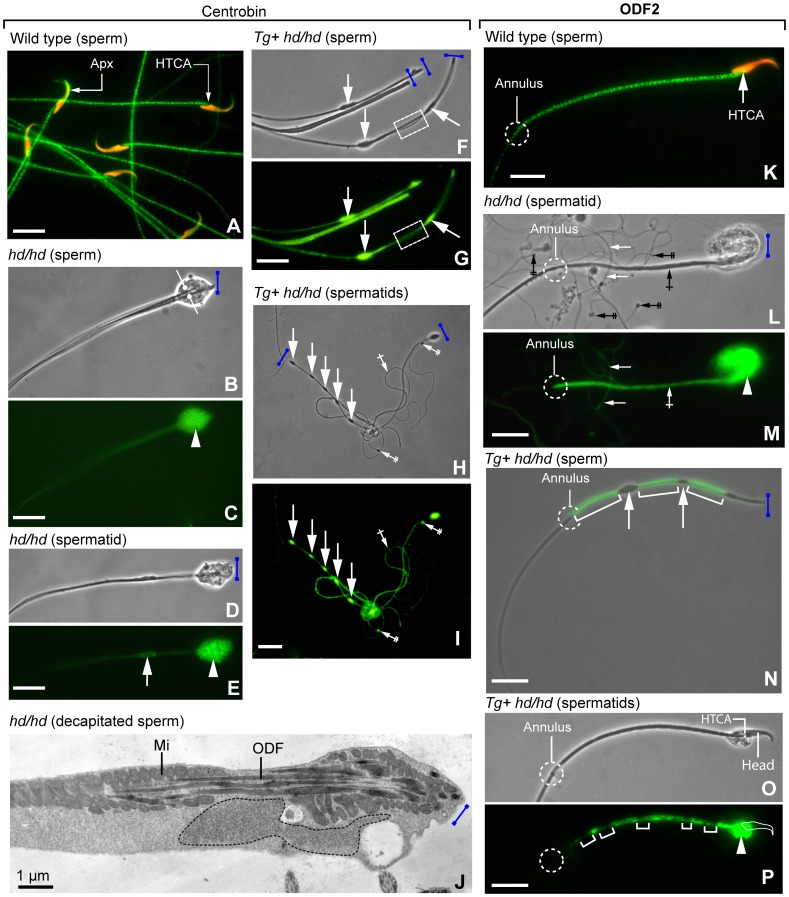
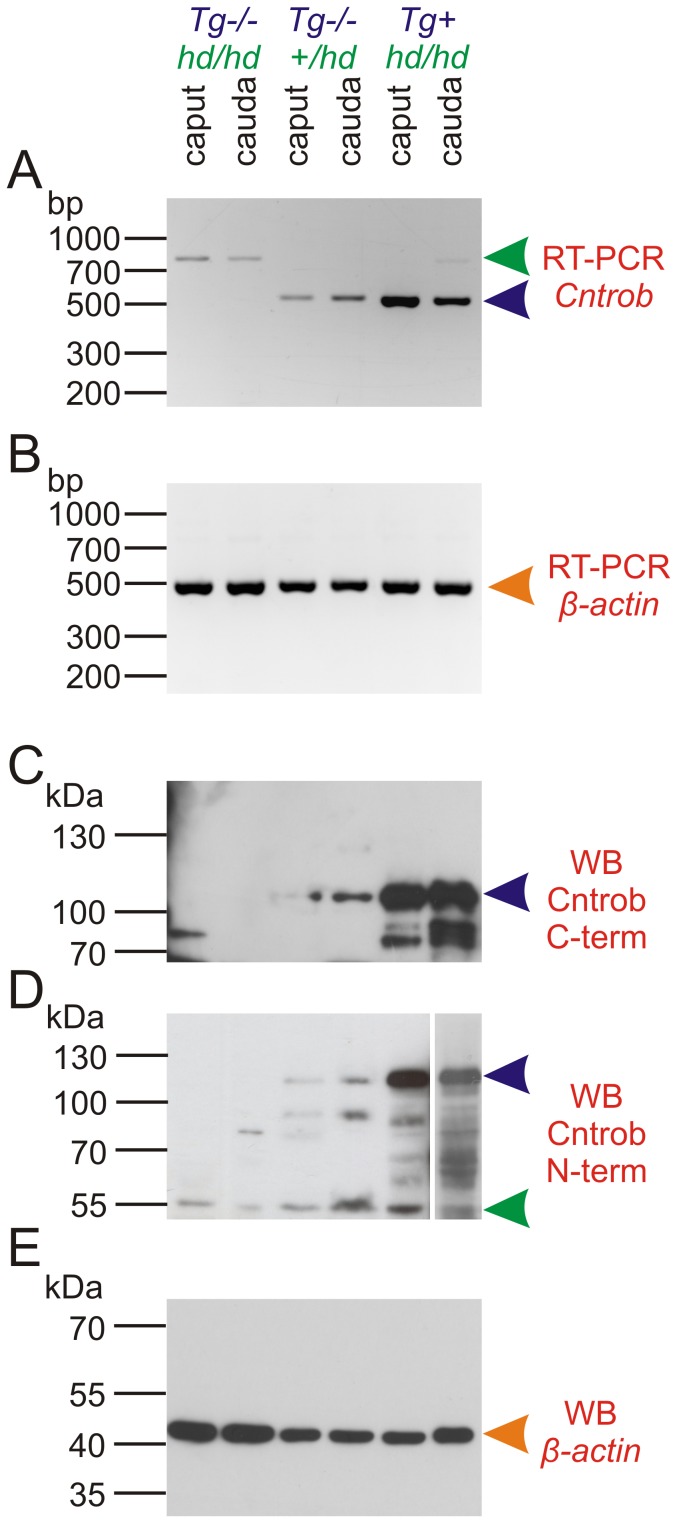
Rat hypodactyly (hd) mutation is characterized by abnormal spermatogenesis and sperm decapitation, limb malformation (missing digits II and III) and growth retardation. We have previously reported centrobin (centrosome BRCA2-interacting protein) truncation at the C-terminus in the hd mutant. Here, we report data from a transgenic rescue experiment carried out to determine a role of centrobin in pathogenesis of hd. The transgenic construct, consisting of full-length-coding cDNA linked to a ubiquitous strong promoter/enhancer combination, was inserted to chromosome 16 into a LINE repeat. No known gene is present in the vicinity of the insertion site. Transgenic centrobin was expressed in all tissues tested, including testis. Transgenic animals show normal body weight and limb morphology as well as average weight of testis and epididymis. Yet, abnormal spermatogenesis and sperm decapitation persisted in the transgenic animals. Western blotting showed the coexistence of full-length and truncated or partially degraded centrobin in sperm of the rescued transgenic animals. Immunocytochemistry showed a buildup of centrobin and ODF2 (outer dense fiber 2) at the sperm decapitation site in the hd mutant and rescued transgenic rats. Additional findings included bulge-like formations and thread-like focal dissociations along the sperm flagellum and the organization of multiple whorls of truncated sperm flagella in the epididymal lumen. We conclude that centrobin is essential for normal patterning of the limb autopod. Centrobin may be required for stabilizing the attachment of the sperm head to flagellum and for maintaining the structural integrity of the sperm flagellum. We postulate that the presence of truncated centrobin, coexisting with full-length centrobin, together with incorrect timing of transgenic centrobin expression may hamper the rescue of fertility in hd male rats.
Conflict of interest statement
Figures
References
-
- Boivin J, Bunting L, Collins JA, Nygren KG (2007) International estimates of infertility prevalence and treatment-seeking: potential need and demand for infertility medical care. Hum Reprod 22: 1506–1512. - PubMed
-
- Liška F (2003) Selected genetic aspects of male infertility–what animal models tell us. Folia Biol (Praha) 49: 129–141. - PubMed
-
- Manandhar G, Simerly C, Schatten G (2000) Centrosome reduction during mammalian spermiogenesis. Curr Top Dev Biol 49: 343–363. - PubMed
-
- Woolley DM, Fawcett DW (1973) The degeneration and disappearance of the centrioles during the development of the rat spermatozoon. Anat Rec 177: 289–301. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
