

Stepwise assembly of the human replicative polymerase holoenzyme
- PMID: 23577232
- PMCID: PMC3614016
- DOI: 10.7554/eLife.00278
Stepwise assembly of the human replicative polymerase holoenzyme
Abstract
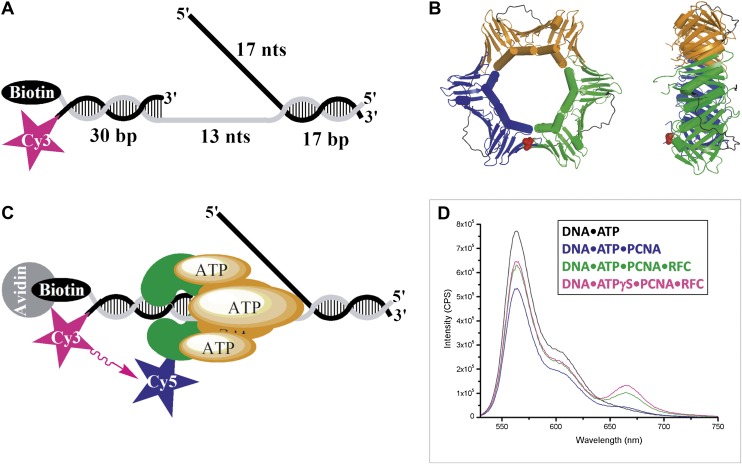
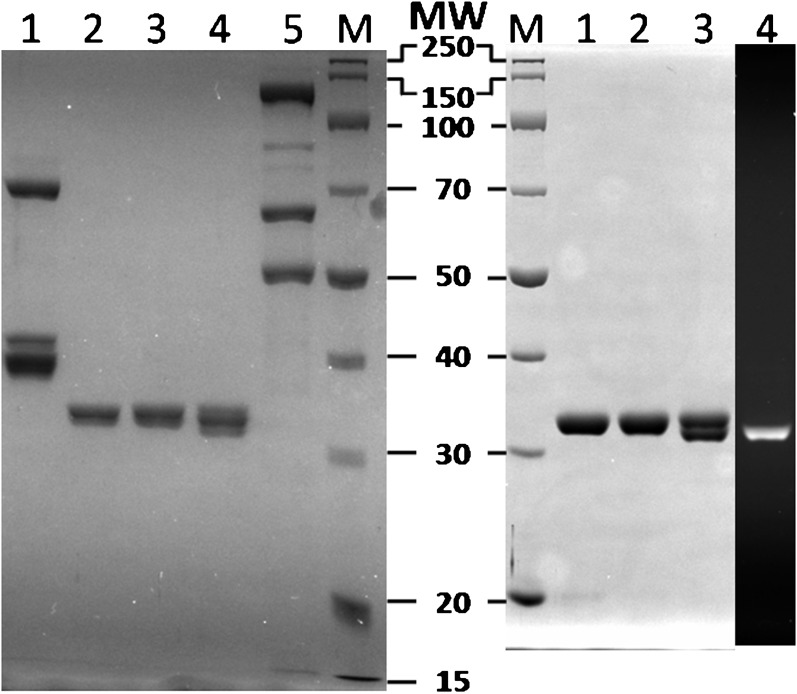
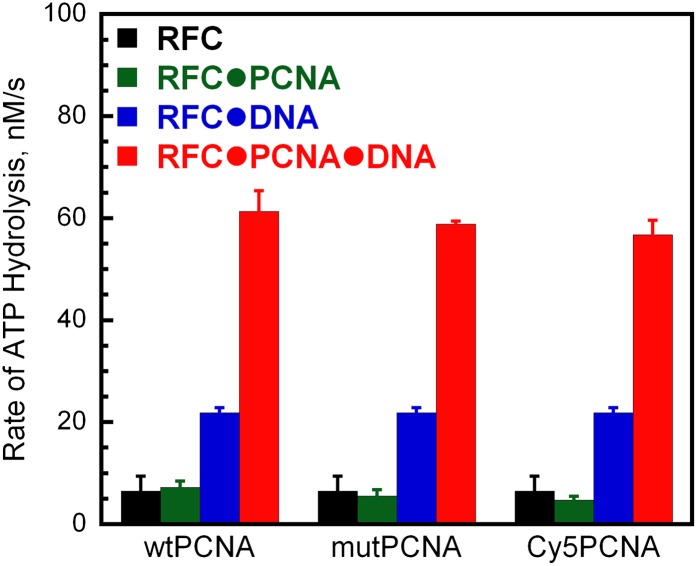
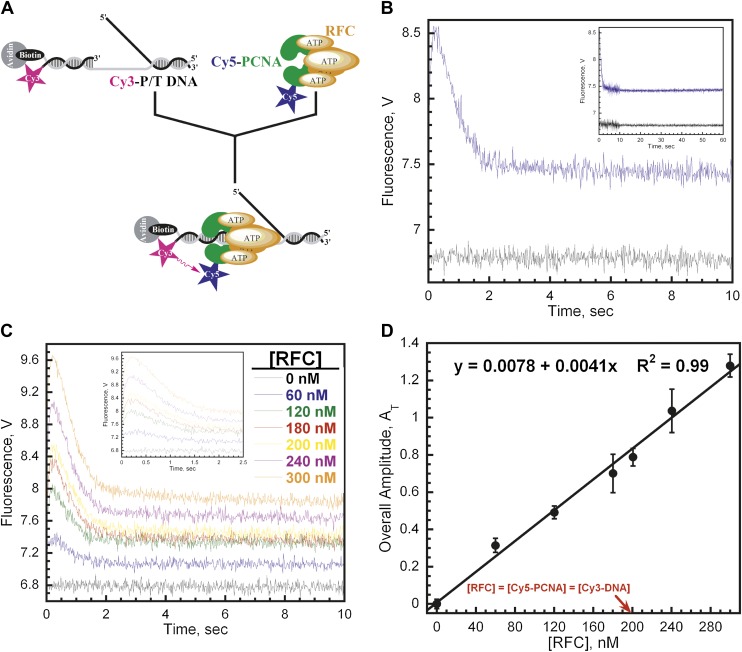
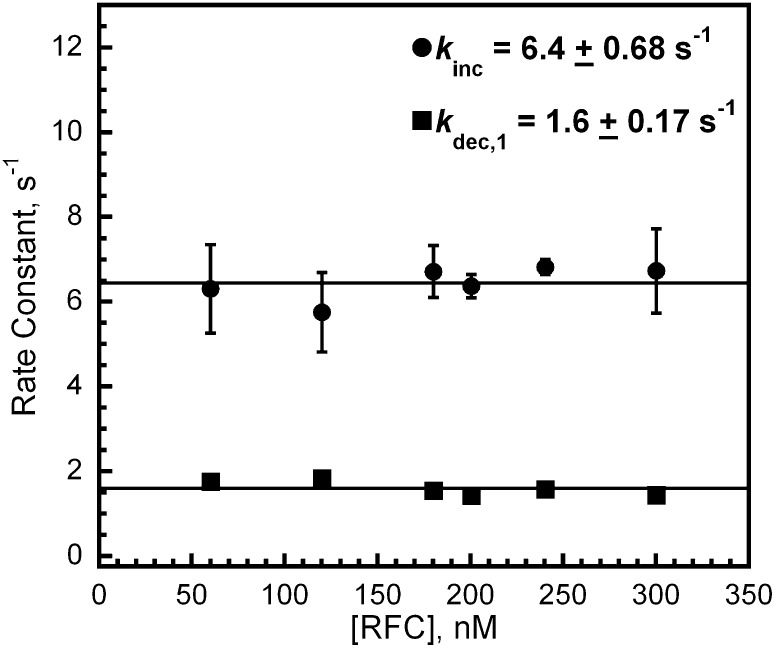
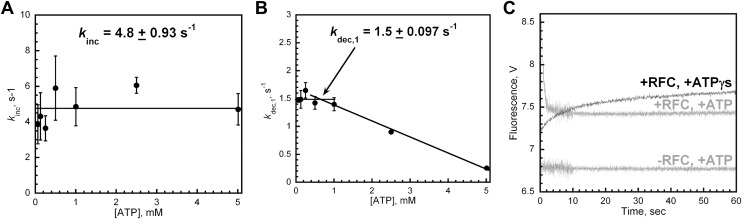
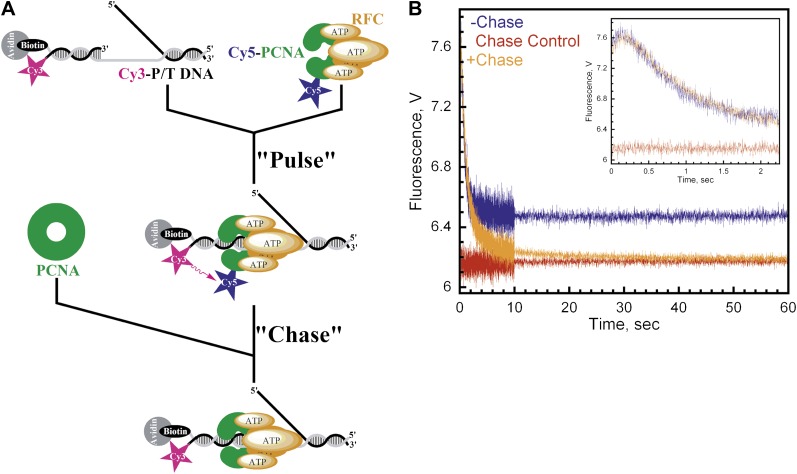
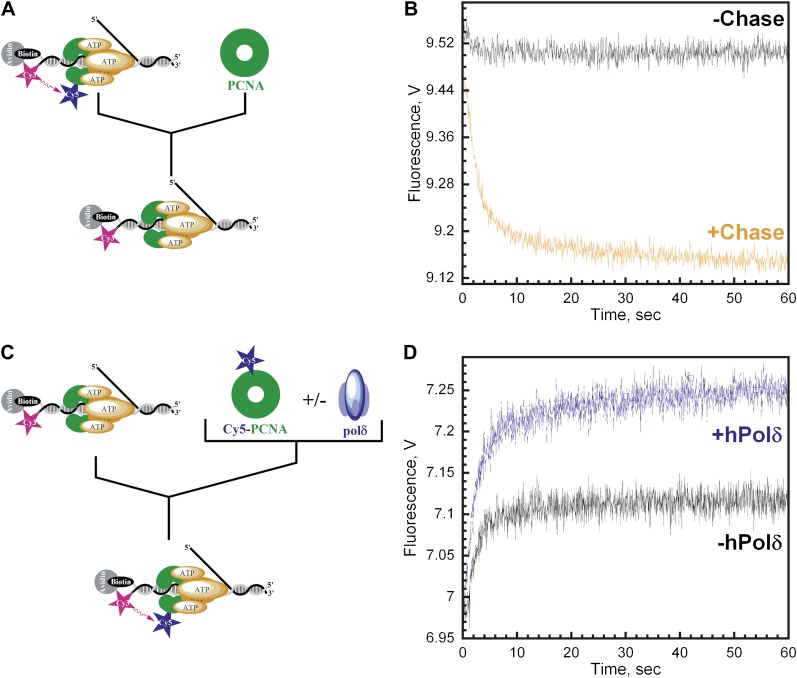
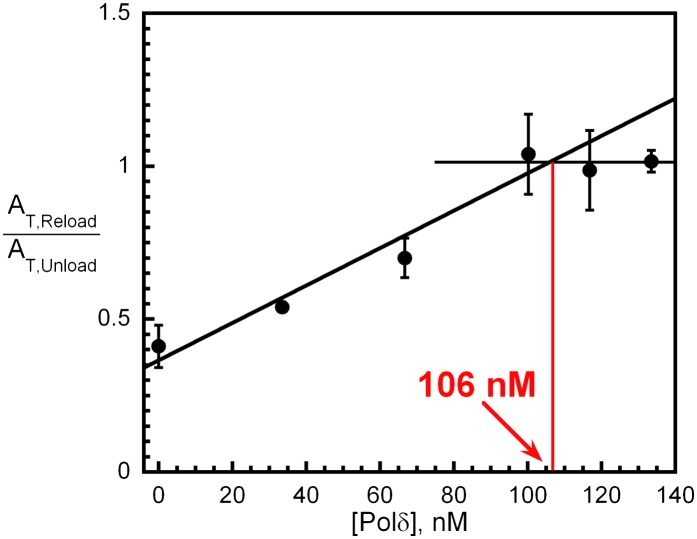
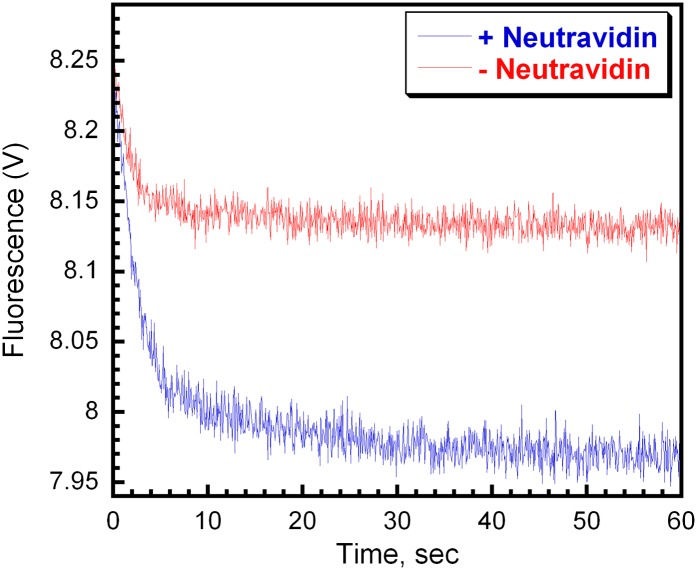
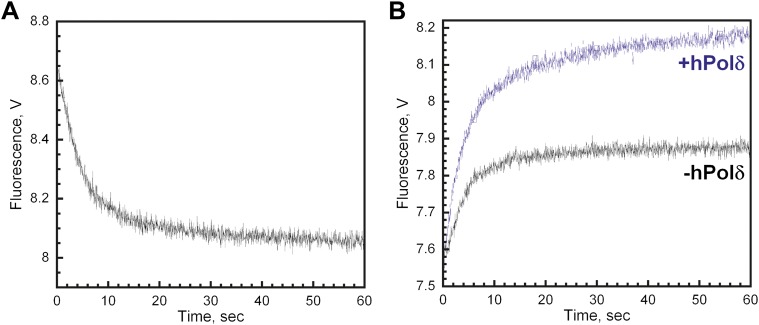
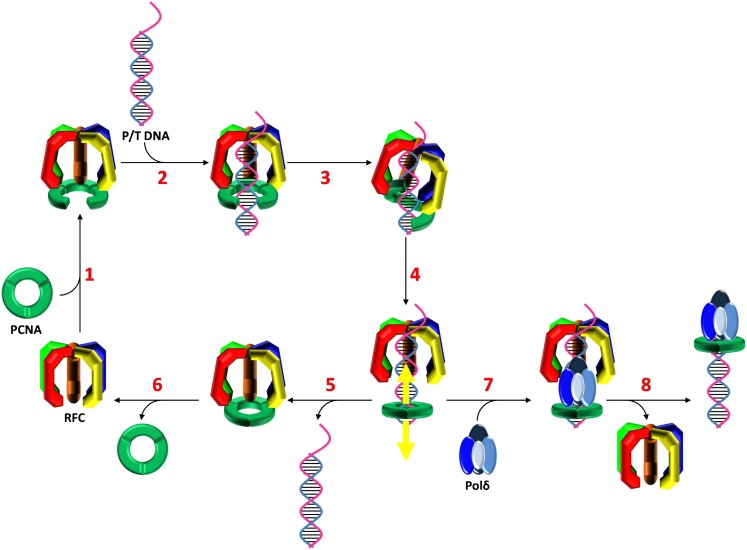
In most organisms, clamp loaders catalyze both the loading of sliding clamps onto DNA and their removal. How these opposing activities are regulated during assembly of the DNA polymerase holoenzyme remains unknown. By utilizing FRET to monitor protein-DNA interactions, we examined assembly of the human holoenzyme. The results indicate that assembly proceeds in a stepwise manner. The clamp loader (RFC) loads a sliding clamp (PCNA) onto a primer/template junction but remains transiently bound to the DNA. Unable to slide away, PCNA re-engages with RFC and is unloaded. In the presence of polymerase (polδ), loaded PCNA is captured from DNA-bound RFC which subsequently dissociates, leaving behind the holoenzyme. These studies suggest that the unloading activity of RFC maximizes the utilization of PCNA by inhibiting the build-up of free PCNA on DNA in the absence of polymerase and recycling limited PCNA to keep up with ongoing replication. DOI:http://dx.doi.org/10.7554/eLife.00278.001.
Keywords: DNA polymerase holoenzyme assembly; Human; clamp loader; clamp loading; clamp unloading; sliding clamp.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures

References
-
- Cai J, Gibbs E, Uhlmann F, Phillips B, Yao N, O'Donnell M, et al. 1997. A complex consisting of human replication factor C p40, p37, and p36 subunits is a DNA-dependent ATPase and an intermediate in the assembly of the holoenzyme. J Biol Chem 272:18974–81 doi: 10.1074/jbc.272.30.18974 - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
