

Principles of cotranslational ubiquitination and quality control at the ribosome
- PMID: 23583075
- PMCID: PMC3886275
- DOI: 10.1016/j.molcel.2013.03.010
Principles of cotranslational ubiquitination and quality control at the ribosome
Abstract
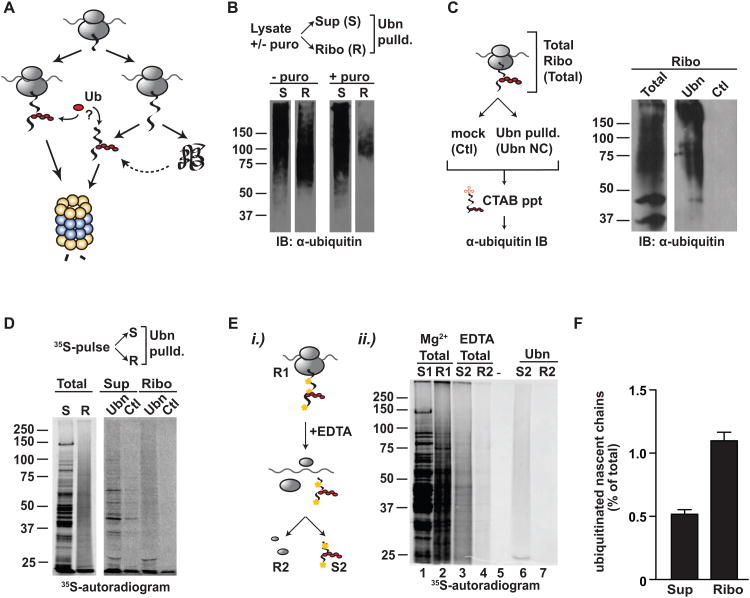
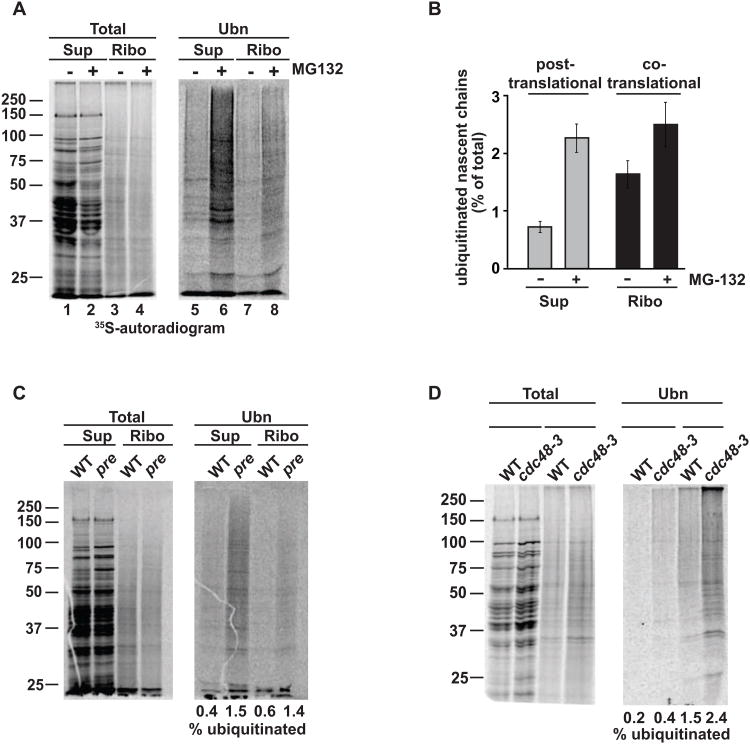
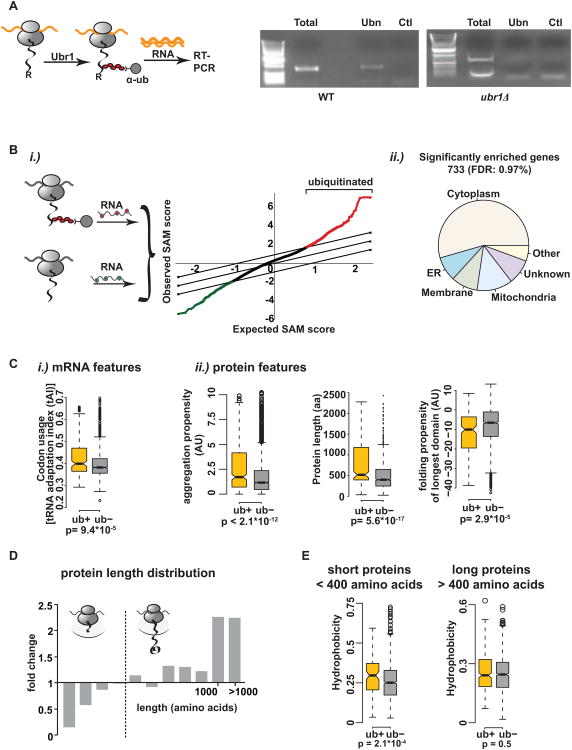
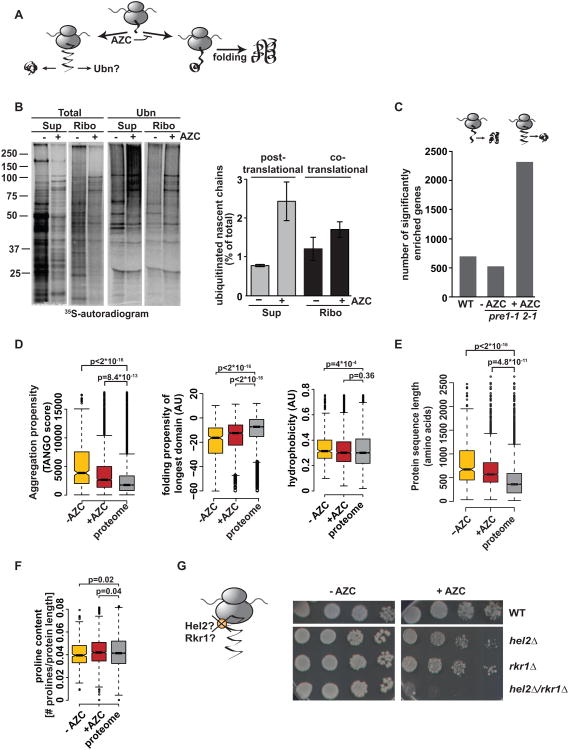
Achieving efficient cotranslational folding of complex proteomes poses a challenge for eukaryotic cells. Nascent polypeptides that emerge vectorially from the ribosome often cannot fold stably and may be susceptible to misfolding and degradation. The extent to which nascent chains are subject to cotranslational quality control and degradation remains unclear. Here, we directly and quantitatively assess cotranslational ubiquitination and identify, at a systems level, the determinants and factors governing this process. Cotranslational ubiquitination occurs at very low levels and is carried out by a complex network of E3 ubiquitin ligases. Ribosome-associated chaperones and cotranslational folding protect the majority of nascent chains from premature quality control. Nonetheless, a number of nascent chains whose intrinsic properties hinder efficient cotranslational folding remain susceptible for cotranslational ubiquitination. We find that quality control at the ribosome is achieved through a tiered system wherein nascent polypeptides have a chance to fold before becoming accessible to ubiquitination.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures


Comment in
-
Protein metabolism: Quality control at the ribosome.Nat Rev Mol Cell Biol. 2013 Jun;14(6):323. doi: 10.1038/nrm3583. Epub 2013 May 2. Nat Rev Mol Cell Biol. 2013. PMID: 23636497 No abstract available.
References
-
- Albanese V, Yam AY, Baughman J, Parnot C, Frydman J. Systems analyses reveal two chaperone networks with distinct functions in eukaryotic cells. Cell. 2006;124:75–88. - PubMed
-
- Balch WE, Morimoto RI, Dillin A, Kelly JW. Science. Vol. 319. New York, NY: 2008. Adapting proteostasis for disease intervention; pp. 916–919. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
