

Variation in the dorsal gradient distribution is a source for modified scaling of germ layers in Drosophila
- PMID: 23583556
- PMCID: PMC3731453
- DOI: 10.1016/j.cub.2013.03.031
Variation in the dorsal gradient distribution is a source for modified scaling of germ layers in Drosophila
Abstract
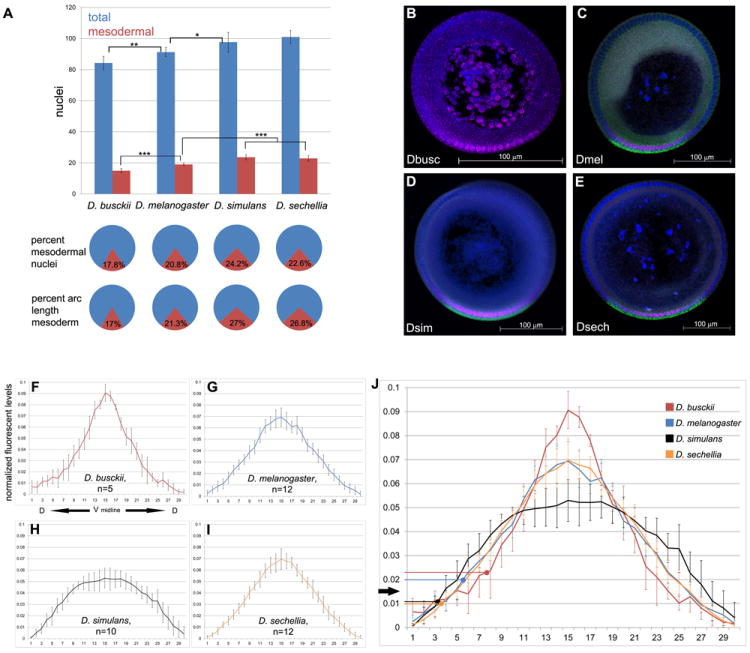
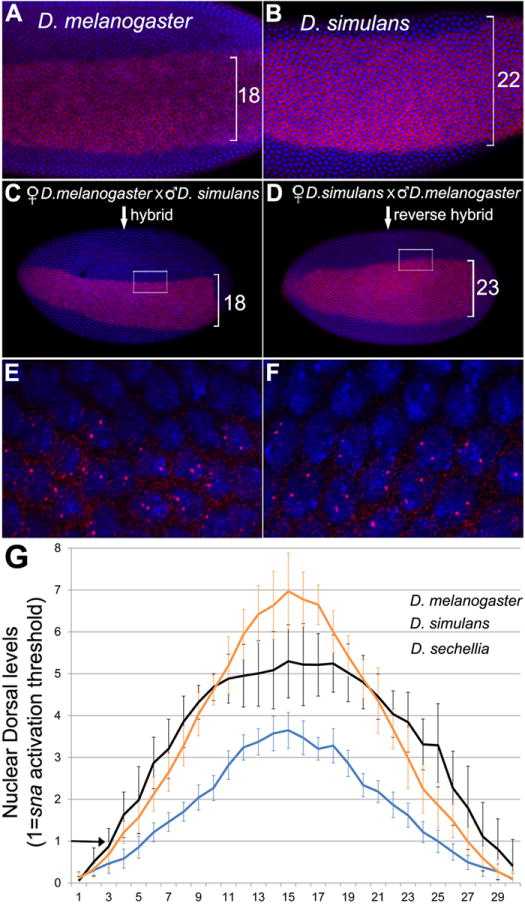
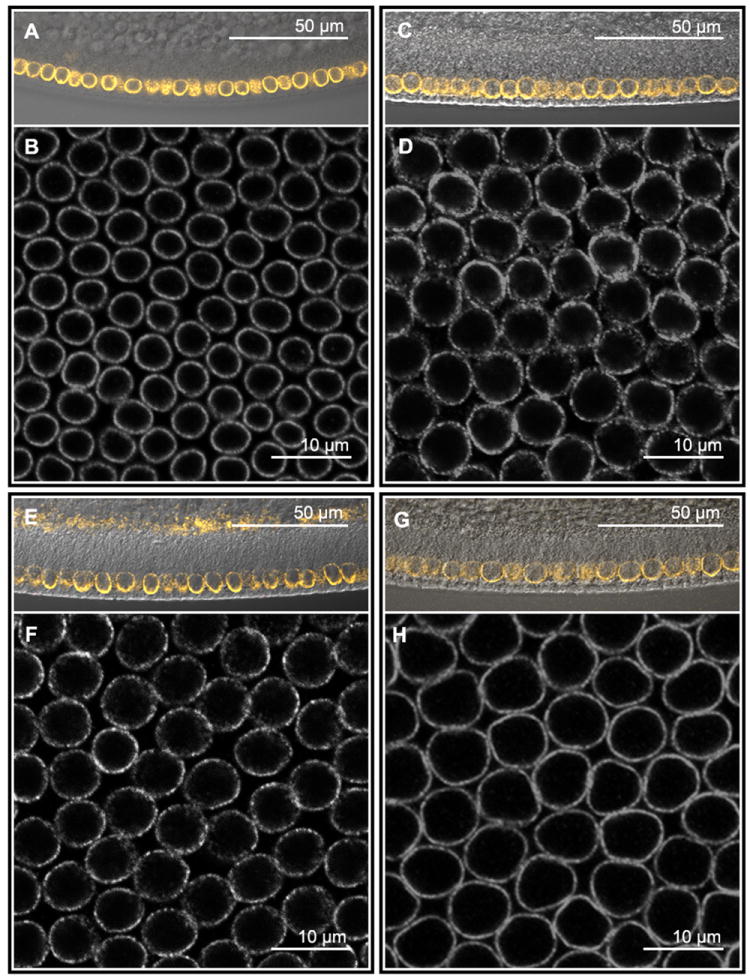
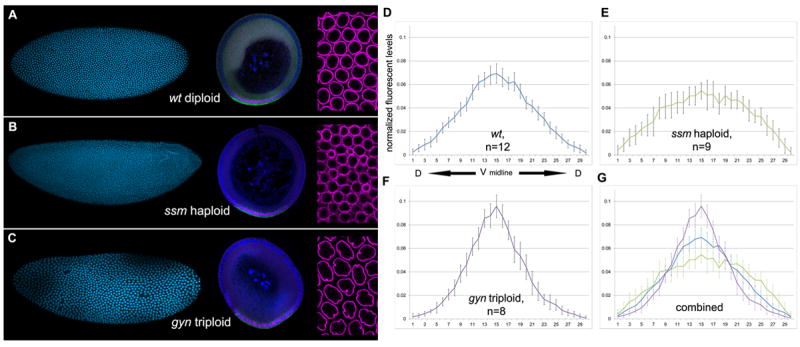
Specification of germ layers along the dorsoventral axis by morphogenetic gradients is an ideal model to study scaling properties of gradients and cell fate changes during evolution. Classical anatomical studies in divergent insects (e.g., flies and grasshoppers) revealed that the neuroectodermal size is conserved and originates similar numbers of neuroblasts of homologous identity. In contrast, mesodermal domains vary significantly in closely related Drosophila species. To further investigate the underlying mechanisms of scaling of germ layers across Drosophila species, we quantified the Dorsal (Dl)/NF-κB gradient, the main morphogenetic gradient that initiates separation of the mesoderm, neuroectoderm, and ectoderm. We discovered a variable range of Toll activation across species and found that Dl activates mesodermal genes at the same threshold levels in melanogaster sibling species. We also show that the Dl gradient distribution can be modulated by nuclear size and packing densities. We propose that variation in mesodermal size occurs at a fast evolutionary rate and is an important mechanism to define the ventral boundary of the neuroectoderm.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures
References
-
- Doe CQ. Molecular markers for identified neuroblasts and ganglion mother cells in the Drosophila central nervous system. Development. 1992;116:855–863. - PubMed
-
- Thomas JB, Bastiani MJ, Bate M, Goodman CS. From grasshopper to Drosophila: a common plan for neuronal development. Nature. 1984;310:203–207. - PubMed
-
- Whitington P. Evolution of neural development in the arthropods. Seminars in Cell & Developmental Biology. 1996;7:605–614.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
