

A unified model for left-right asymmetry? Comparison and synthesis of molecular models of embryonic laterality
- PMID: 23583583
- PMCID: PMC3698617
- DOI: 10.1016/j.ydbio.2013.03.021
A unified model for left-right asymmetry? Comparison and synthesis of molecular models of embryonic laterality
Abstract
Understanding how and when the left-right (LR) axis is first established is a fundamental question in developmental biology. A popular model is that the LR axis is established relatively late in embryogenesis, due to the movement of motile cilia and the resultant directed fluid flow during late gastrulation/early neurulation. Yet, a large body of evidence suggests that biophysical, molecular, and bioelectrical asymmetries exist much earlier in development, some as early as the first cell cleavage after fertilization. Alternative models of LR asymmetry have been proposed that accommodate these data, postulating that asymmetry is established due to a chiral cytoskeleton and/or the asymmetric segregation of chromatids. There are some similarities, and many differences, in how these various models postulate the origin and timing of symmetry breaking and amplification, and these events' linkage to the well-conserved subsequent asymmetric transcriptional cascades. This review examines experimental data that lend strong support to an early origin of LR asymmetry, yet are also consistent with later roles for cilia in the amplification of LR pathways. In this way, we propose that the various models of asymmetry can be unified: early events are needed to initiate LR asymmetry, and later events could be utilized by some species to maintain LR-biases. We also present an alternative hypothesis, which proposes that individual embryos stochastically choose one of several possible pathways with which to establish their LR axis. These two hypotheses are both tractable in appropriate model species; testing them to resolve open questions in the field of LR patterning will reveal interesting new biology of wide relevance to developmental, cell, and evolutionary biology.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
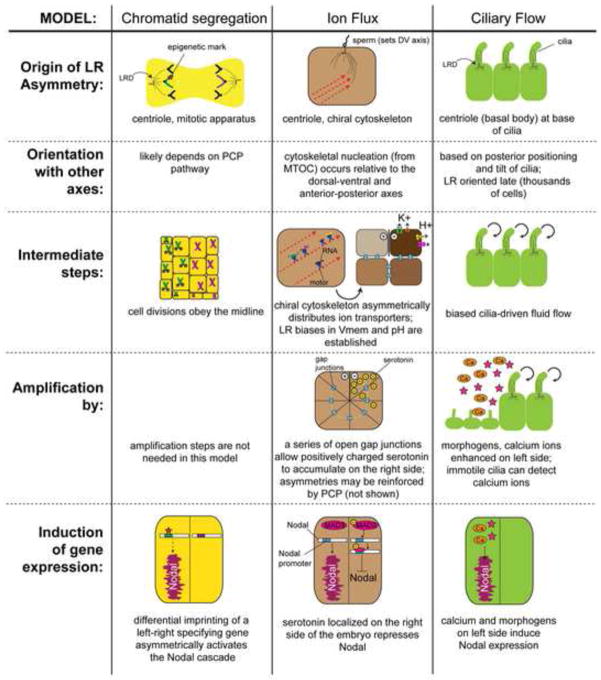
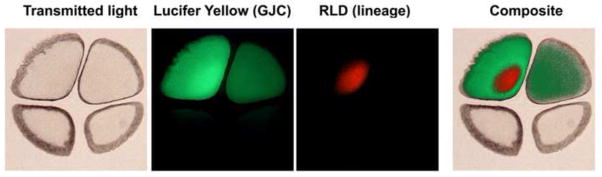
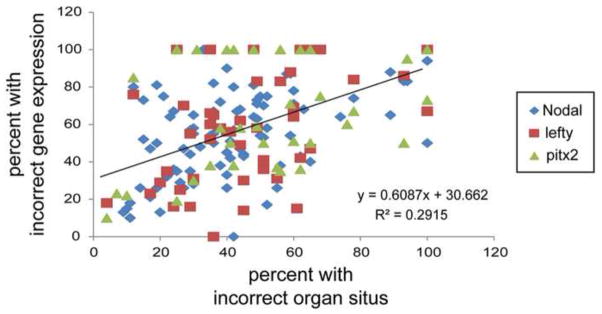
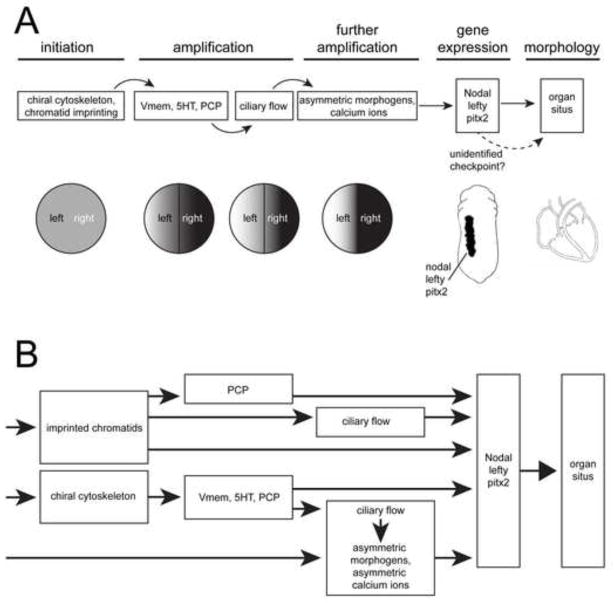
References
-
- Abe T, Thitamadee S, Hashimoto T. Microtubule defects and cell morphogenesis in the lefty1lefty2 tubulin mutant of Arabidopsis thaliana. Plant Cell Physiol. 2004;45:211–220. - PubMed
-
- Adler PN, Taylor J, Charlton J. The domineering non-autonomy of frizzled and van Gogh clones in the Drosophila wing is a consequence of a disruption in local signaling. Mech Dev. 2000;96:197–207. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
