

Macrophage ADAM17 deficiency augments CD36-dependent apoptotic cell uptake and the linked anti-inflammatory phenotype
- PMID: 23584255
- PMCID: PMC3693752
- DOI: 10.1161/CIRCRESAHA.112.300683
Macrophage ADAM17 deficiency augments CD36-dependent apoptotic cell uptake and the linked anti-inflammatory phenotype
Abstract
Rationale: Apoptotic cell phagocytosis (efferocytosis) is mediated by specific receptors and is essential for resolution of inflammation. In chronic inflammation, apoptotic cell clearance is dysfunctional and soluble levels of several apoptotic cell receptors are elevated. Reports have identified proteolytic cleavage as a mechanism capable of releasing soluble apoptotic cell receptors, but the functional implications of their proteolysis are unclear.
Objective: To test the hypothesis that ADAM17-mediated cleavage of apoptotic cell receptors limits efferocytosis in vivo.
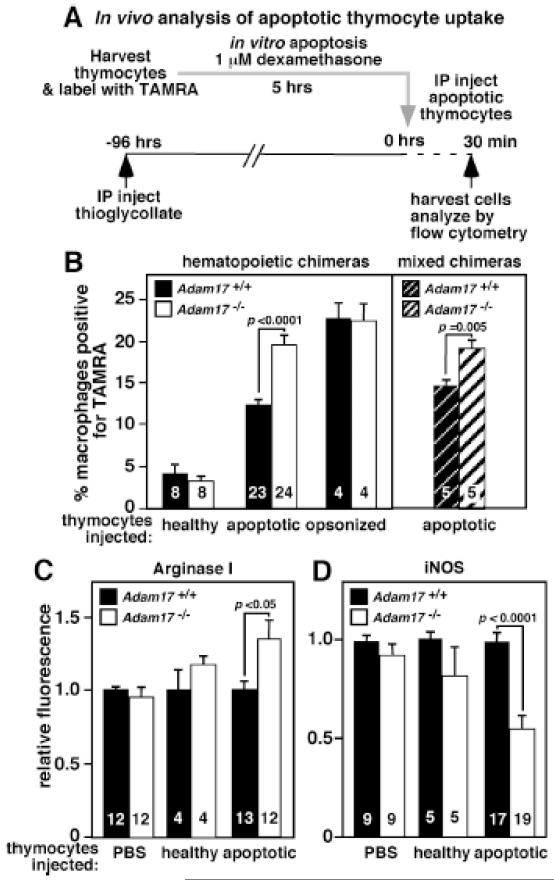
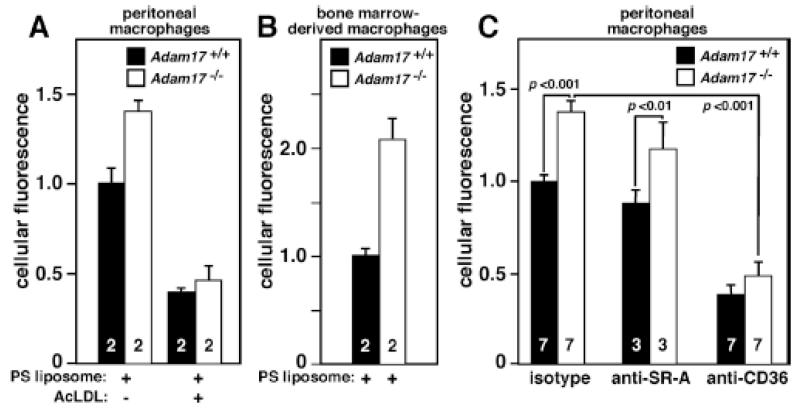
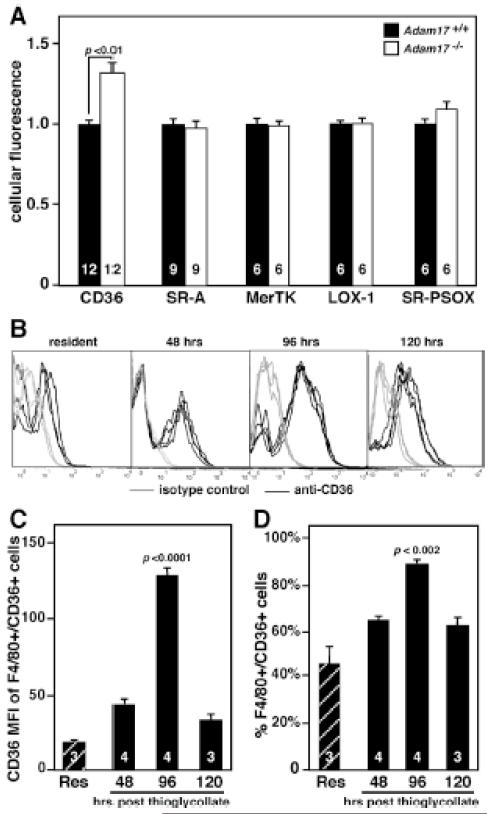
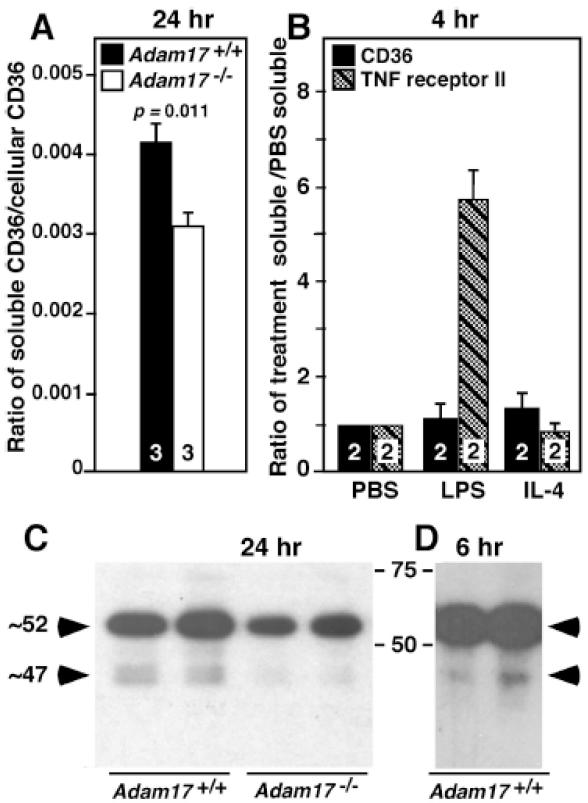
Methods and results: In vivo comparison of macrophage efferocytosis in wild-type and Adam17-null hematopoietic chimeras demonstrates that ADAM17 deficiency leads to a 60% increase in efferocytosis and an enhanced anti-inflammatory phenotype in a model of peritonitis. In vitro uptake of phosphatidylserine liposomes identifies the dual-pass apoptotic cell receptor CD36 as a major contributor to enhanced efferocytosis, and CD36 surface levels are elevated on macrophages from Adam17-null mice. Further, temporal elevation of CD36 expression with inflammation may also contribute to its impact. Soluble CD36 from macrophage-conditioned media comprises 2 species based on Western blotting, and mass spectrometry identifies 3 N-terminal peptides that represent probable cleavage sites. Levels of soluble CD36 are decreased in Adam17-null conditioned media, providing evidence for involvement of ADAM17 in CD36 cleavage. Importantly, enhanced efferocytosis in vivo by macrophages lacking ADAM17 is CD36 dependent and accelerates macrophage clearance from the peritoneum, thus promoting resolution of inflammation and highlighting the impact of increased apoptotic cell uptake.
Conclusions: Our studies demonstrate the importance of ADAM17-mediated proteolysis for in vivo efferocytosis regulation and suggest a possible mechanistic link between chronic inflammation and defective efferocytosis.
Keywords: apoptotic cells; inflammation; macrophage; metalloproteinase; proteolysis.
Figures
Comment in
-
Shedding light on impaired efferocytosis and nonresolving inflammation.Circ Res. 2013 Jun 21;113(1):9-12. doi: 10.1161/CIRCRESAHA.113.301583. Circ Res. 2013. PMID: 23788501 Free PMC article.
References
-
- Devitt A, Marshall LJ. The innate immune system and the clearance of apoptotic cells. J Leukoc Biol. 2011;90:447–457. - PubMed
-
- Schrijvers DM, De Meyer GR, Kockx MM, Herman AG, Martinet W. Phagocytosis of apoptotic cells by macrophages is impaired in atherosclerosis. Arterioscler Thromb Vasc Biol. 2005;25:1256–1261. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
