

Substrate selectivity of the acid-activated glutamate/γ-aminobutyric acid (GABA) antiporter GadC from Escherichia coli
- PMID: 23589309
- PMCID: PMC3663535
- DOI: 10.1074/jbc.M113.474502
Substrate selectivity of the acid-activated glutamate/γ-aminobutyric acid (GABA) antiporter GadC from Escherichia coli
Abstract
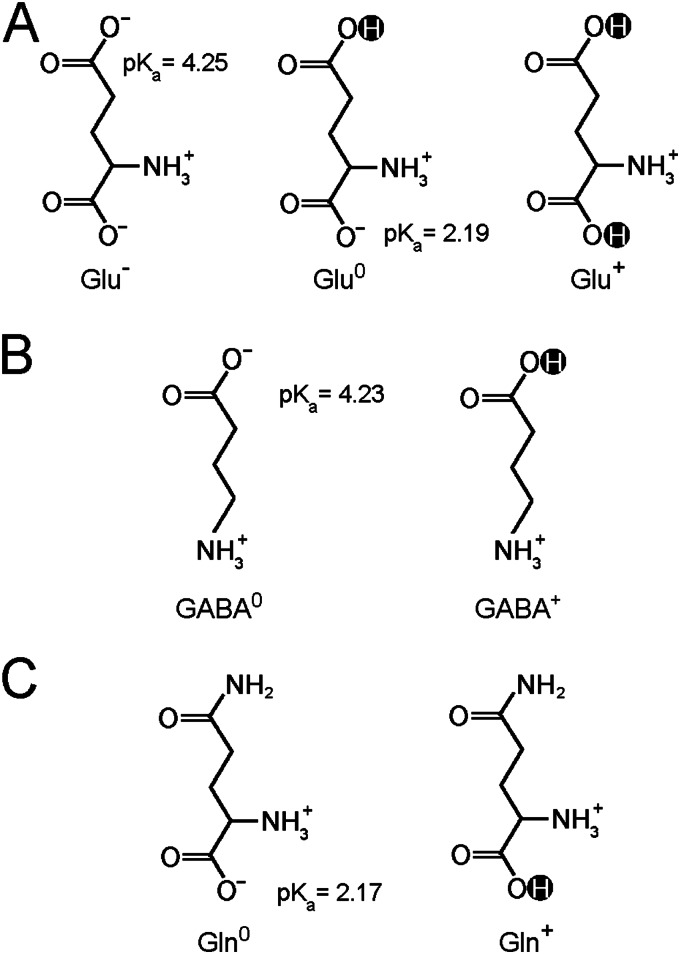
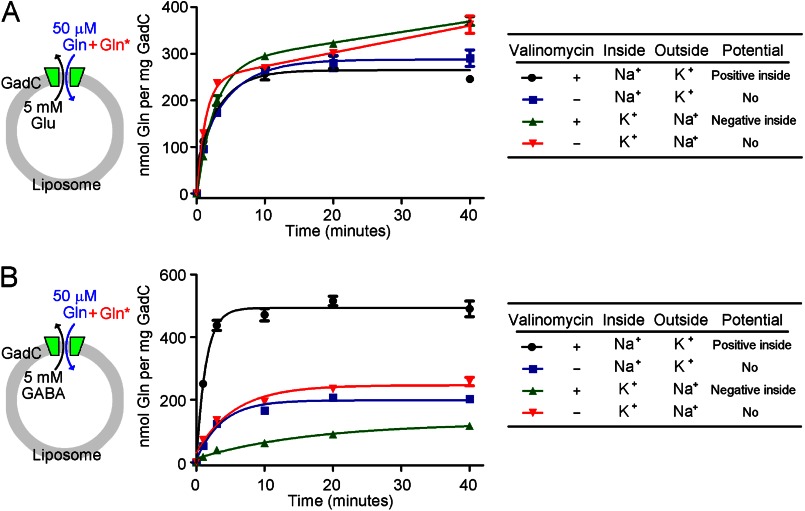
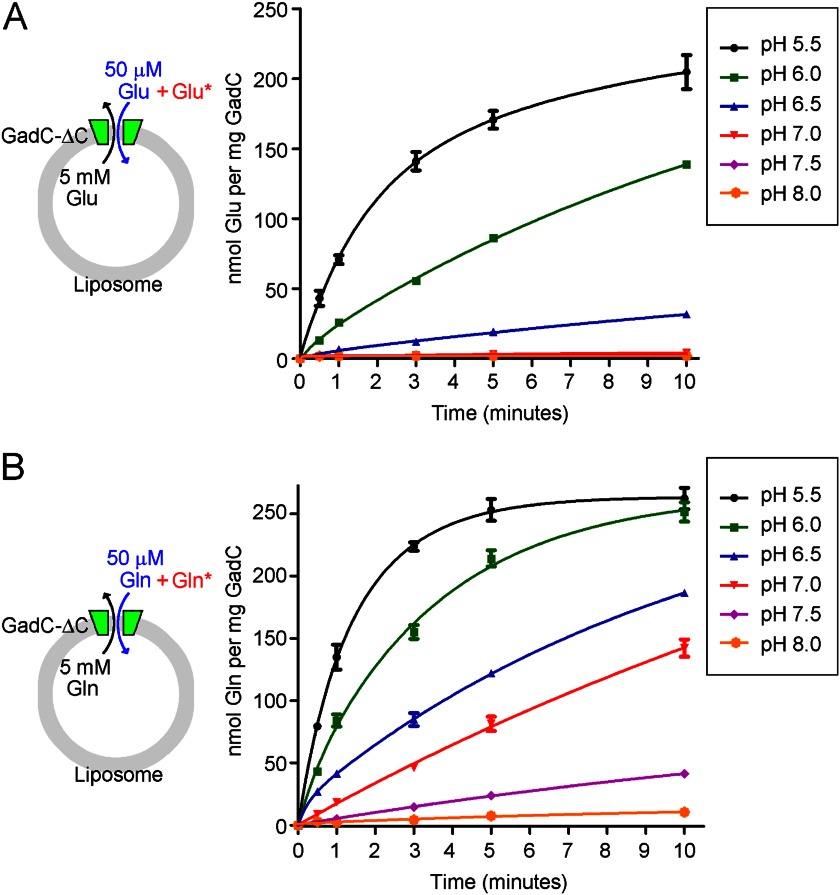
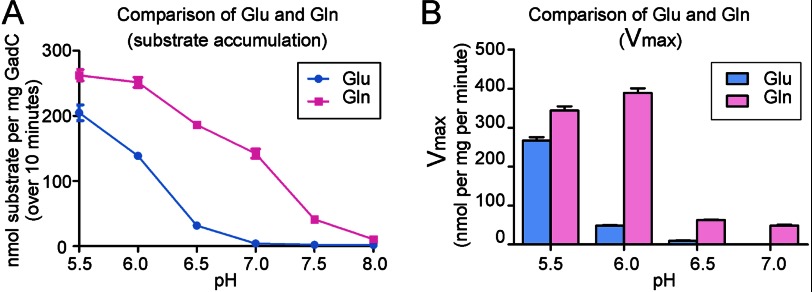
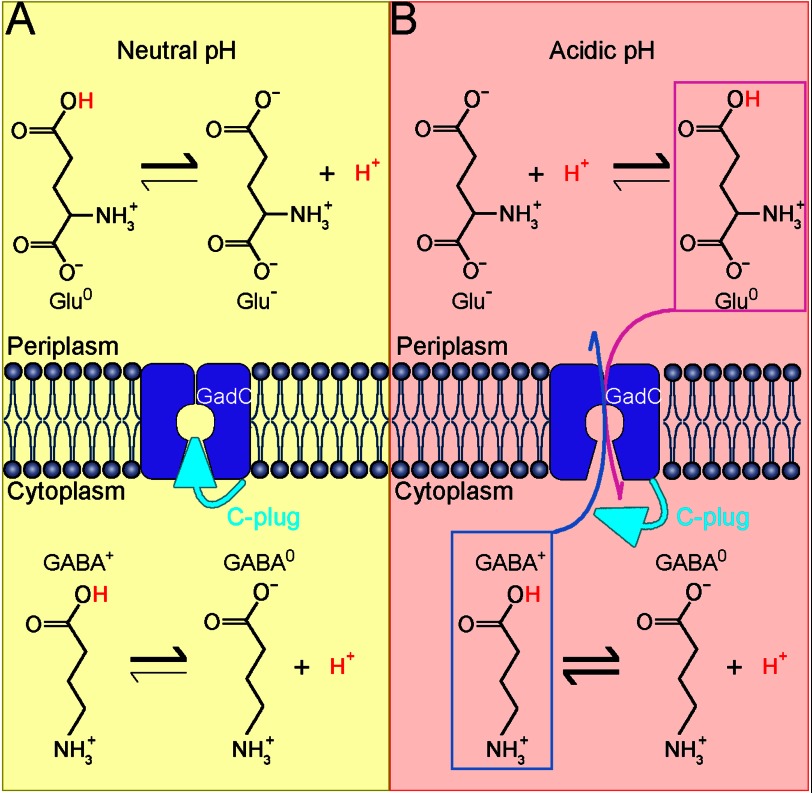
GadC, a central component of the Escherichia coli acid resistance system, is a Glu/GABA antiporter. A previous structural study and biochemical characterization showed that GadC exhibits a stringent pH dependence for substrate transport, with no detectable activity at pH values above 6.5. However, the substrate selectivity and the mechanism of pH-dependent transport activity of GadC remain enigmatic. In this study, we demonstrate that GadC selectively transports Glu with no net charge and GABA with a positive charge. A C-plug-truncated variant of GadC (residues 1-470) transported Gln (a mimic of Glu with no net charge), but not Glu, even at pH 8.0. The pH-dependent transport of Gln by this GadC variant was shifted ~1 unit toward a higher pH compared with Glu transport. Taken together, the results identify the substrate selectivity for GadC and show that the protonation states of substrates are crucial for transport.
Keywords: Acid Resistance; Amino Acid Transport; GadC; Glutamate; Glutamine; Membrane Antiporter; Membrane Proteins; Membrane Transporter Reconstitution; Protonation/Deprotonation; Substrate Selectivity.
Figures
References
-
- Donnenberg M. S. (2000) Pathogenic strategies of enteric bacteria. Nature 406, 768–774 - PubMed
-
- Audia J. P., Webb C. C., Foster J. W. (2001) Breaking through the acid barrier: an orchestrated response to proton stress by enteric bacteria. Int. J. Med. Microbiol. 291, 97–106 - PubMed
-
- Foster J. W. (2004) Escherichia coli acid resistance: tales of an amateur acidophile. Nat. Rev. Microbiol. 2, 898–907 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
