

Phosphoregulation promotes release of kinetochores from dynamic microtubules via multiple mechanisms
- PMID: 23589891
- PMCID: PMC3645574
- DOI: 10.1073/pnas.1220700110
Phosphoregulation promotes release of kinetochores from dynamic microtubules via multiple mechanisms
Abstract
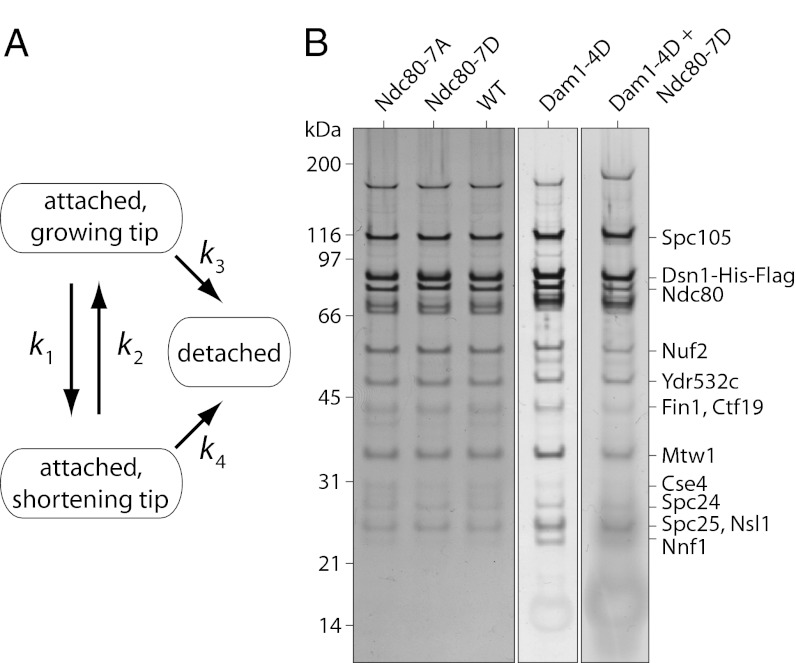
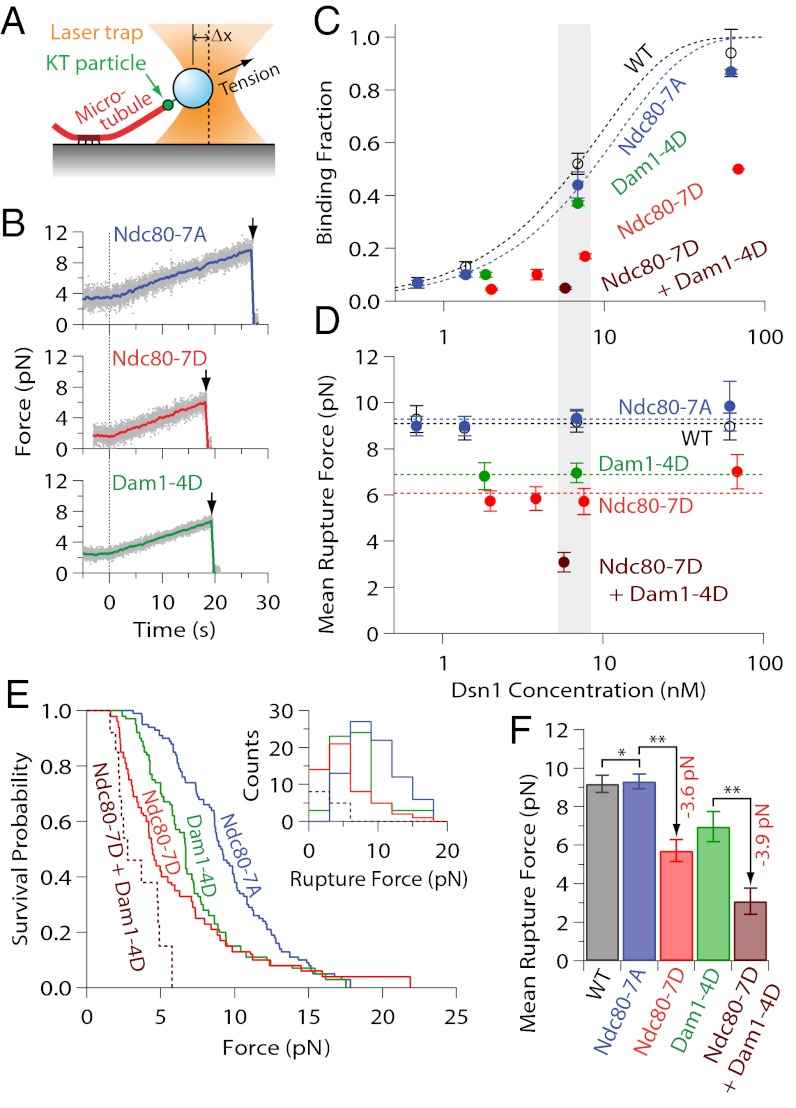
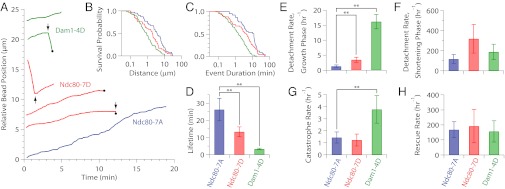
During mitosis, multiprotein complexes called kinetochores orchestrate chromosome segregation by forming load-bearing attachments to dynamic microtubule tips, and by participating in phosphoregulatory error correction. The conserved kinase Aurora B phosphorylates the major microtubule-binding kinetochore subcomplexes, Ndc80 and (in yeast) Dam1, to promote release of erroneous attachments, giving another chance for proper attachments to form. It is unknown whether Aurora B phosphorylation promotes release directly, by increasing the rate of kinetochore detachment, or indirectly, by destabilizing the microtubule tip. Moreover, the relative importance of phosphorylation of Ndc80 vs. Dam1 in the context of whole kinetochores is unclear. To address these uncertainties, we isolated native yeast kinetochore particles carrying phosphomimetic mutations on Ndc80 and Dam1, and applied advanced laser-trapping techniques to measure the strength and stability of their attachments to individual dynamic microtubule tips. Rupture forces were reduced by phosphomimetic mutations on both subcomplexes, in an additive manner, indicating that both subcomplexes make independent contributions to attachment strength. Phosphomimetics on either subcomplex reduced attachment lifetimes under constant force, primarily by accelerating detachment during microtubule growth. Phosphomimetics on Dam1 also increased the likelihood of switches from microtubule growth into shortening, further promoting release in an indirect manner. Taken together, our results suggest that, in vivo, Aurora B releases kinetochores via at least two mechanisms: by weakening the kinetochore-microtubule interface and also by destabilizing the kinetochore-attached microtubule tip.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
