

Dynamic switching of calmodulin interactions underlies Ca2+ regulation of CaV1.3 channels
- PMID: 23591884
- PMCID: PMC3856249
- DOI: 10.1038/ncomms2727
Dynamic switching of calmodulin interactions underlies Ca2+ regulation of CaV1.3 channels
Abstract
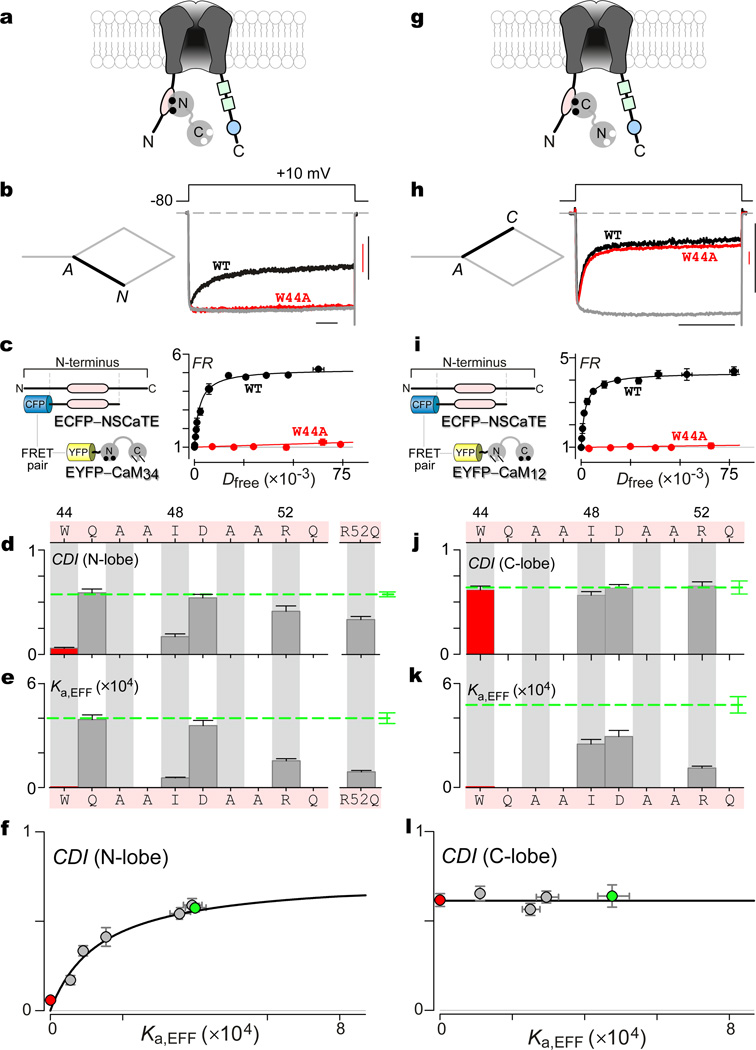
Calmodulin regulation of CaV channels is a prominent Ca(2+) feedback mechanism orchestrating vital adjustments of Ca(2+) entry. The long-held structural correlation of this regulation has been Ca(2+)-bound calmodulin, complexed alone with an IQ domain on the channel carboxy terminus. Here, however, systematic alanine mutagenesis of the entire carboxyl tail of an L-type CaV1.3 channel casts doubt on this paradigm. To identify the actual molecular states underlying channel regulation, we develop a structure-function approach relating the strength of regulation to the affinity of underlying calmodulin/channel interactions, by a Langmuir relation (individually transformed Langmuir analysis). Accordingly, we uncover frank exchange of Ca(2+)-calmodulin to interfaces beyond the IQ domain, initiating substantial rearrangements of the calmodulin/channel complex. The N-lobe of Ca(2+)-calmodulin binds an N-terminal spatial Ca(2+) transforming element module on the channel amino terminus, whereas the C-lobe binds an EF-hand region upstream of the IQ domain. This system of structural plasticity furnishes a next-generation blueprint for CaV channel modulation.
Figures







References
-
- Halling DB, Aracena-Parks P, Hamilton SL. Regulation of voltage-gated Ca2+ channels by calmodulin. Sci STKE. 2006;2006 er1. - PubMed
-
- Erickson MG, Alseikhan BA, Peterson BZ, Yue DT. Preassociation of calmodulin with voltage-gated Ca(2+) channels revealed by FRET in single living cells. Neuron. 2001;31:973–985. - PubMed
-
- Erickson MG, Liang H, Mori MX, Yue DT. FRET two-hybrid mapping reveals function and location of L-type Ca2+ channel CaM preassociation. Neuron. 2003;39:97–107. - PubMed
-
- Pitt GS, et al. Molecular basis of calmodulin tethering and Ca2+-dependent inactivation of L-type Ca2+ channels. J. Biol. Chem. 2001;276:30794–30802. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
