

The mechanisms of planar cell polarity, growth and the Hippo pathway: some known unknowns
- PMID: 23592229
- PMCID: PMC3677094
- DOI: 10.1016/j.ydbio.2013.01.030
The mechanisms of planar cell polarity, growth and the Hippo pathway: some known unknowns
Abstract
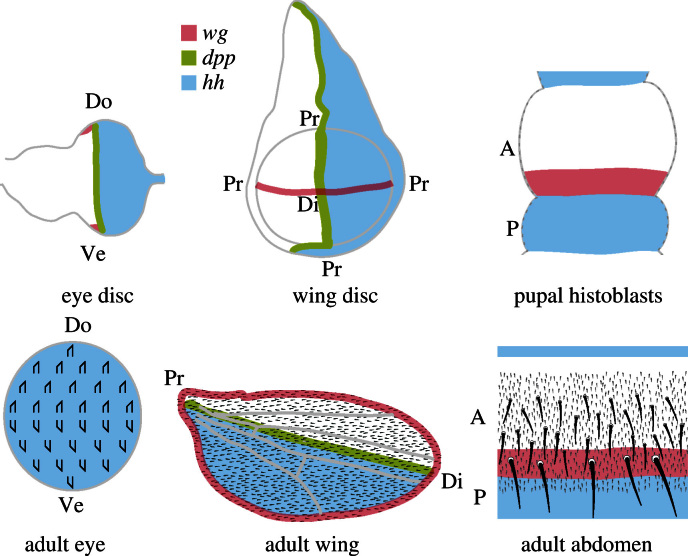
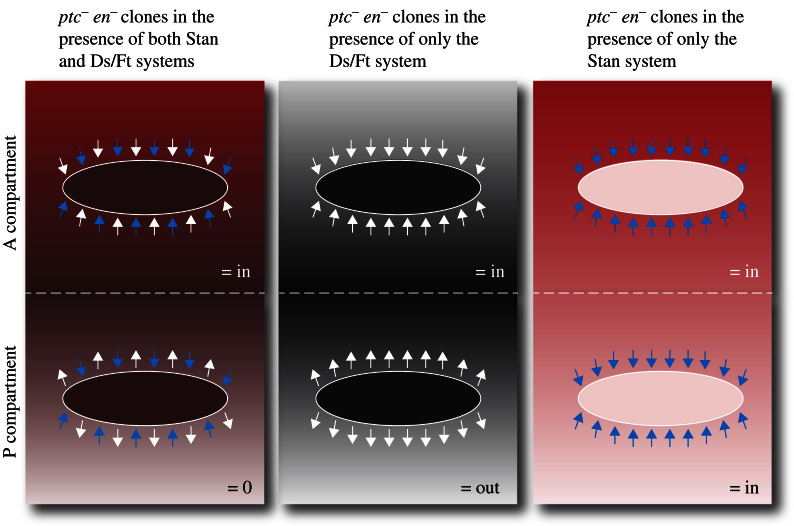
Planar cell polarity (PCP) is a small but important area of research. In this review we discuss a limited number of topics within the PCP field, chosen because they are difficult, unsolved, controversial or just because we find them interesting. Because Drosophila is the best studied and technically most amenable system we have concentrated on it, but also consider some examples from work on vertebrates. Topics discussed include the number of genetic pathways involved in PCP, as well as the causal relationship between embryonic axes, gradients of morphogens and PCP itself. We consider the vexed question of the roles of the Wnt genes in PCP in both vertebrates and Drosophila. We discuss whether the proteins involved in PCP need to be localised asymmetrically in cells in order to function. We criticise the way the Hippo pathway is described in the literature and ask what its wildtype function is. We explore afresh how the Hippo pathway might be linked both to growth and to PCP through the gigantic cadherin molecule Fat. We offer some new ways of making sense of published results, particularly those relating to the Frizzled/Starry night and Dachsous/Fat systems of PCP.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
References
-
- Adler P., Charlton J., Vinson C. Allelic variation at the frizzled locus of Drosophila. Dev. Genet. 1987;8:99–119.
-
- Adler P.N., Charlton J., Liu J. Mutations in the cadherin superfamily member gene dachsous cause a tissue polarity phenotype by altering frizzled signaling. Development. 1998;125:959–968. - PubMed
-
- Adler P.N., Krasnow R.E., Liu J. Tissue polarity points from cells that have higher Frizzled levels towards cells that have lower Frizzled levels. Curr. Biol. 1997;7:940–949. - PubMed
-
- Adler P.N., Taylor J., Charlton J. The domineering non-autonomy of frizzled and van Gogh clones in the Drosophila wing is a consequence of a disruption in local signaling. Mech. Dev. 2000;96:197–207. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
