

Protein trans-splicing of multiple atypical split inteins engineered from natural inteins
- PMID: 23593141
- PMCID: PMC3620165
- DOI: 10.1371/journal.pone.0059516
Protein trans-splicing of multiple atypical split inteins engineered from natural inteins
Abstract
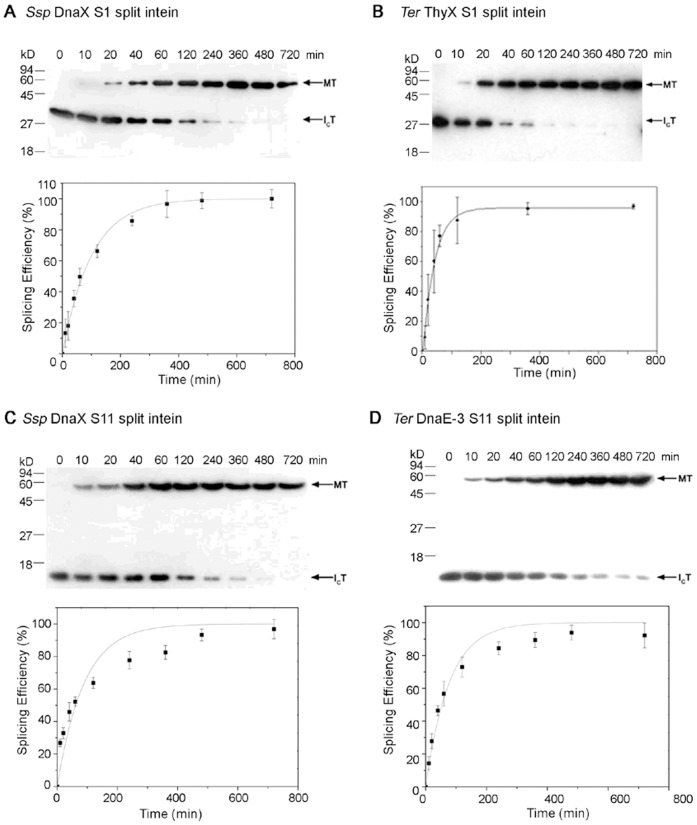
Protein trans-splicing by split inteins has many uses in protein production and research. Splicing proteins with synthetic peptides, which employs atypical split inteins, is particularly useful for site-specific protein modifications and labeling, because the synthetic peptide can be made to contain a variety of unnatural amino acids and chemical modifications. For this purpose, atypical split inteins need to be engineered to have a small N-intein or C-intein fragment that can be more easily included in a synthetic peptide that also contains a small extein to be trans-spliced onto target proteins. Here we have successfully engineered multiple atypical split inteins capable of protein trans-splicing, by modifying and testing more than a dozen natural inteins. These included both S1 split inteins having a very small (11-12 aa) N-intein fragment and S11 split inteins having a very small (6 aa) C-intein fragment. Four of the new S1 and S11 split inteins showed high efficiencies (85-100%) of protein trans-splicing both in E. coli cells and in vitro. Under in vitro conditions, they exhibited reaction rate constants ranging from ~1.7 × 10(-4) s(-1) to ~3.8 × 10(-4) s(-1), which are comparable to or higher than those of previously reported atypical split inteins. These findings should facilitate a more general use of trans-splicing between proteins and synthetic peptides, by expanding the availability of different atypical split inteins. They also have implications on understanding the structure-function relationship of atypical split inteins, particularly in terms of intein fragment complementation.
Conflict of interest statement
Figures

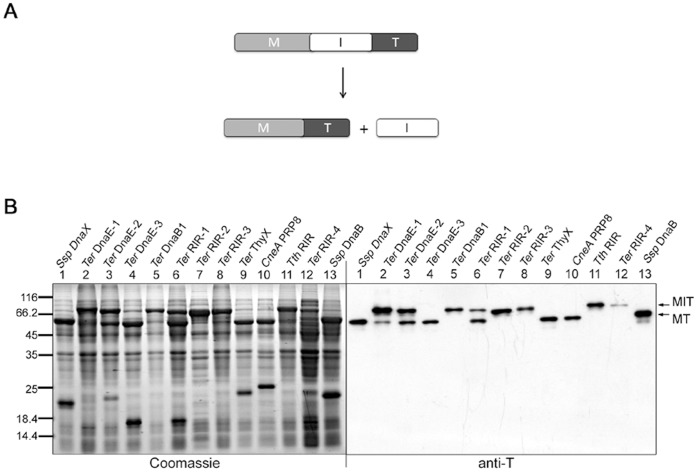
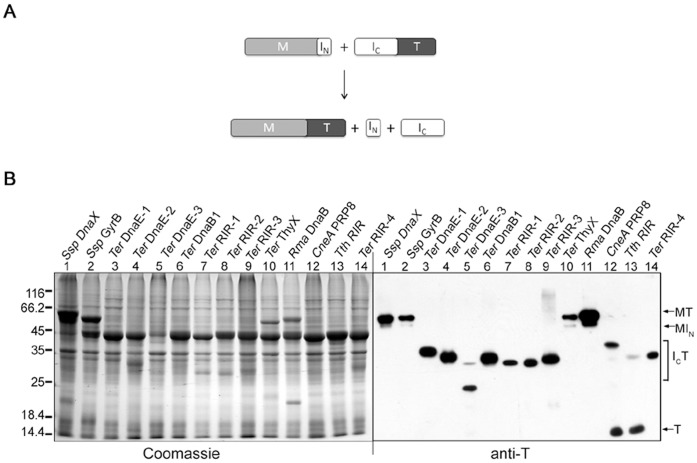
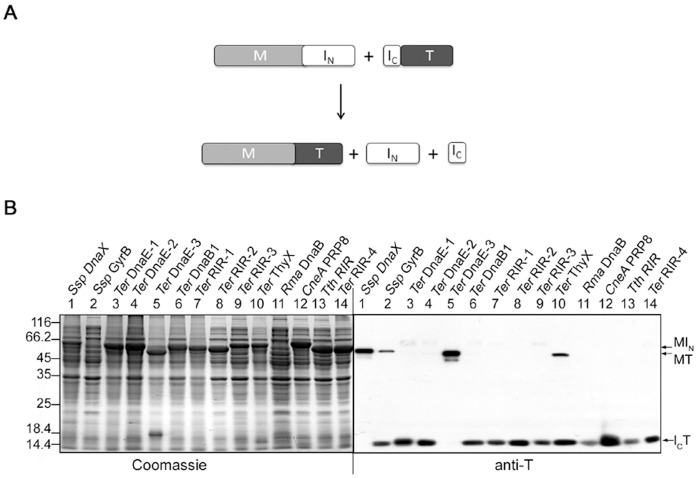
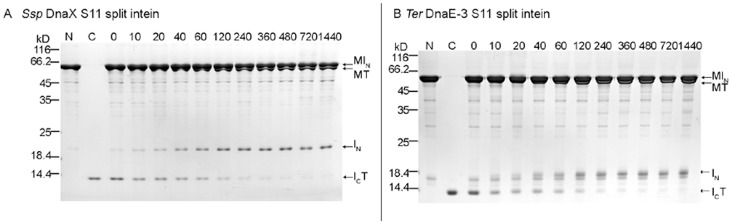
References
-
- Chong S, Shao Y, Paulus H, Benner J, Perler FB, et al. (1996) Protein splicing involving the Saccharomyces cerevisiae VMA intein. The steps in the splicing pathway, side reactions leading to protein cleavage, and establishment of an in vitro splicing system. J Biol Chem 271: 22159–22168. - PubMed
-
- Liu XQ (2000) Protein-splicing intein: Genetic mobility, origin, and evolution. Annu Rev Genet 34: 61–76. - PubMed
-
- Hall TM, Porter JA, Young KE, Koonin EV, Beachy PA, et al. (1997) Crystal structure of a Hedgehog autoprocessing domain: homology between Hedgehog and self-splicing proteins. Cell 91: 85–97. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
