

Willin, an upstream component of the hippo signaling pathway, orchestrates mammalian peripheral nerve fibroblasts
- PMID: 23593160
- PMCID: PMC3620498
- DOI: 10.1371/journal.pone.0060028
Willin, an upstream component of the hippo signaling pathway, orchestrates mammalian peripheral nerve fibroblasts
Abstract
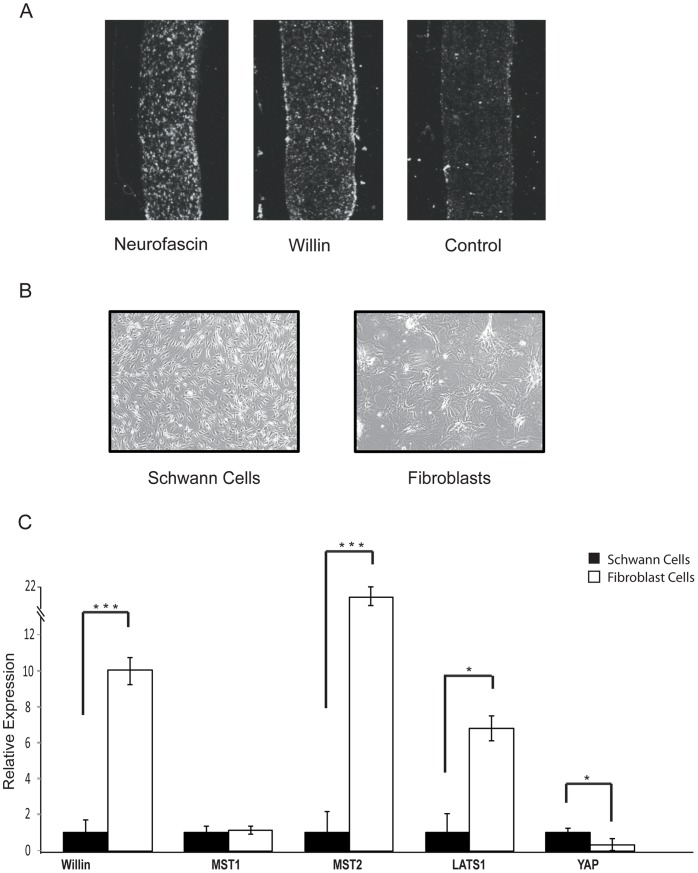
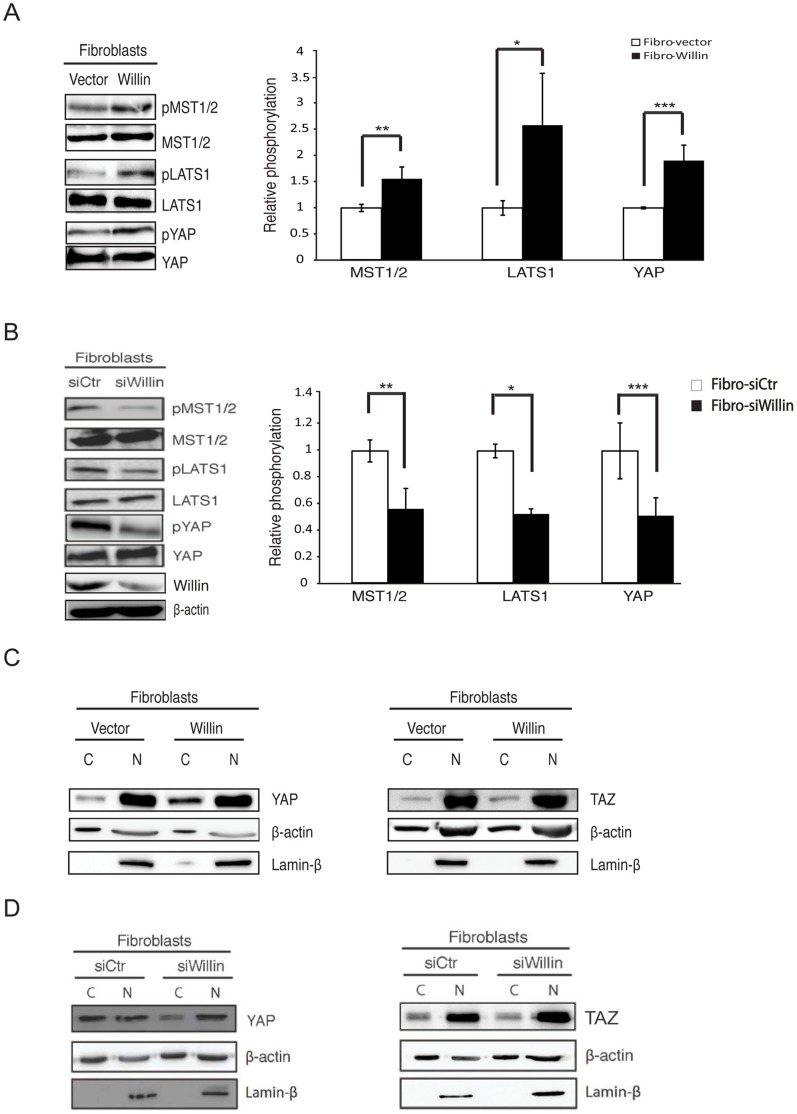
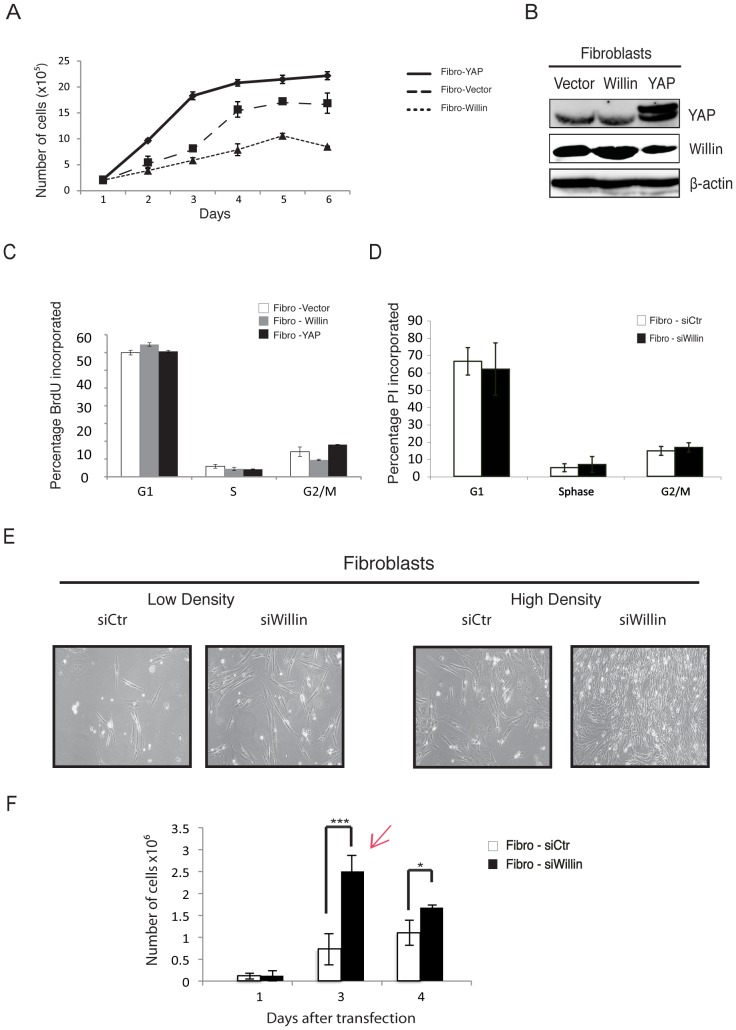
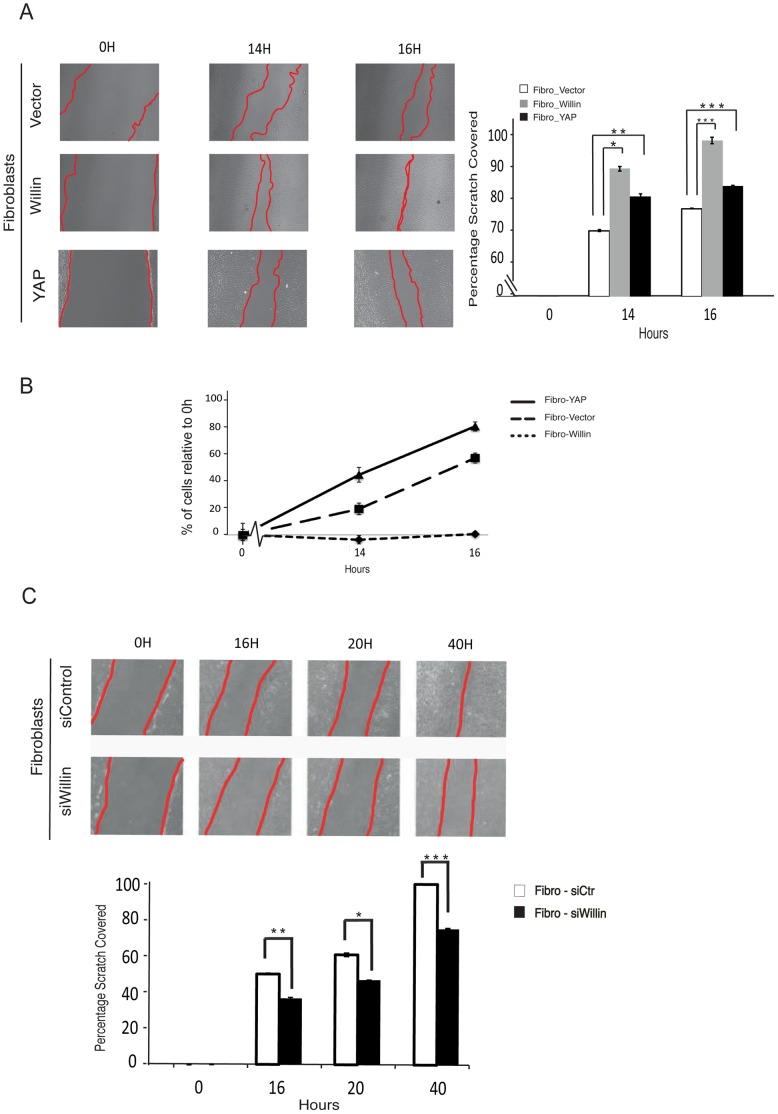
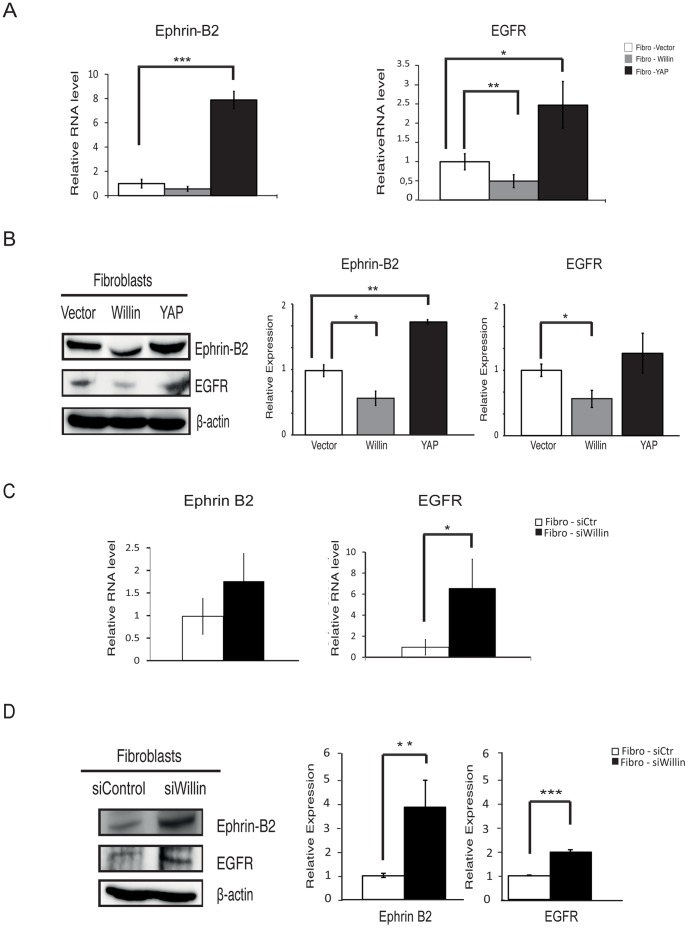
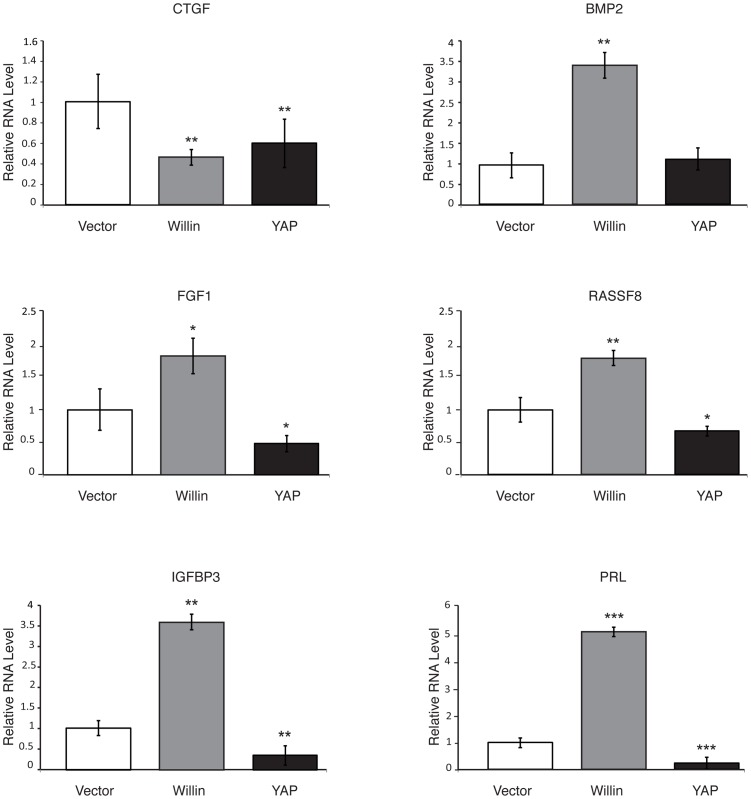
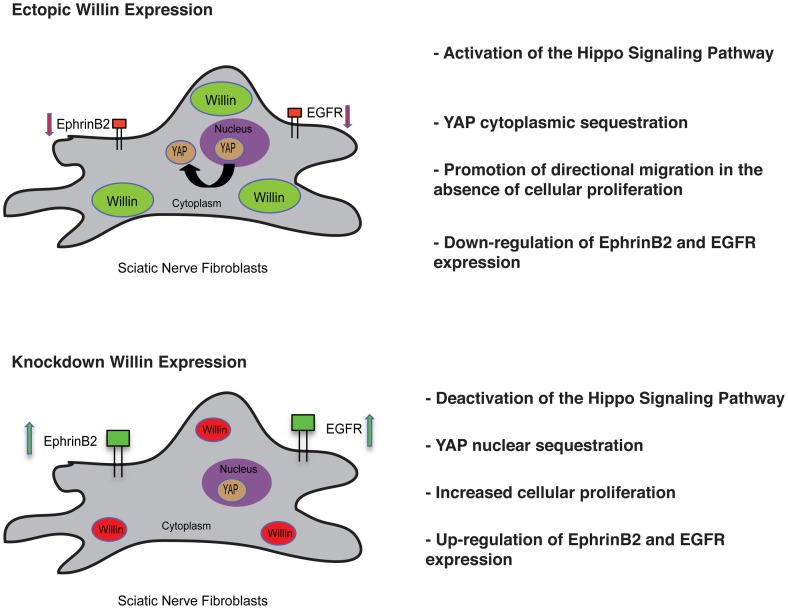
Willin/FRMD6 was first identified in the rat sciatic nerve, which is composed of neurons, Schwann cells, and fibroblasts. Willin is an upstream component of the Hippo signaling pathway, which results in the inactivation of the transcriptional co-activator YAP through Ser127 phosphorylation. This in turn suppresses the expression of genes involved in cell growth, proliferation and cancer development ensuring the control of organ size, cell contact inhibition and apoptosis. Here we show that in the mammalian sciatic nerve, Willin is predominantly expressed in fibroblasts and that Willin expression activates the Hippo signaling cascade and induces YAP translocation from the nucleus to the cytoplasm. In addition within these cells, although it inhibits cellular proliferation, Willin expression induces a quicker directional migration towards scratch closure and an increased expression of factors linked to nerve regeneration. These results show that Willin modulates sciatic nerve fibroblast activity indicating that Willin may have a potential role in the regeneration of the peripheral nervous system.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
