

CD160Ig fusion protein targets a novel costimulatory pathway and prolongs allograft survival
- PMID: 23593209
- PMCID: PMC3617215
- DOI: 10.1371/journal.pone.0060391
CD160Ig fusion protein targets a novel costimulatory pathway and prolongs allograft survival
Abstract
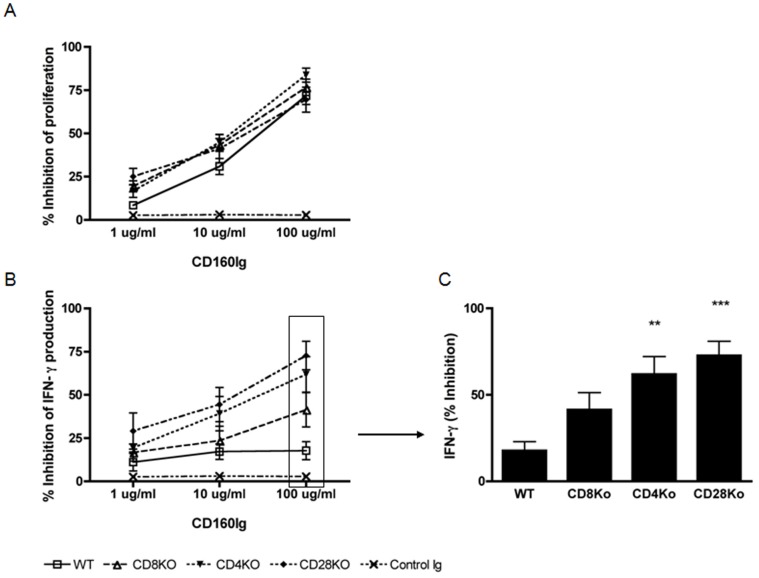
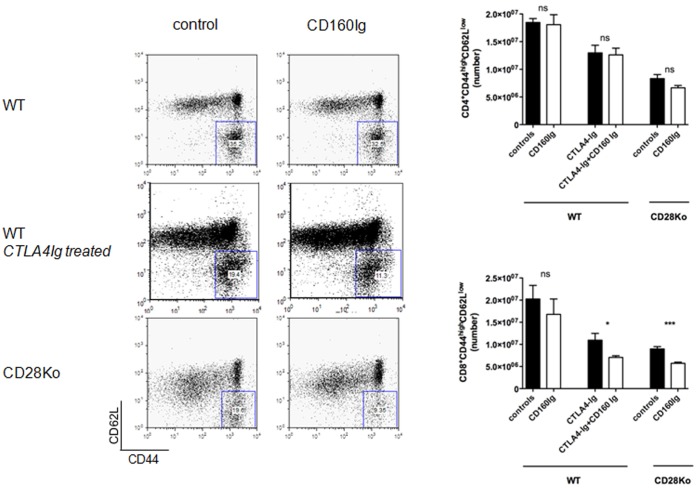
CD160 is a cell surface molecule expressed by most NK cells and approximately 50% of CD8(+) cytotoxic T lymphocytes. Engagement of CD160 by MHC class-I directly triggers a costimulatory signal to TCR-induced proliferation, cytokine production and cytotoxic effector functions. The role of CD160 in alloimmunity is unknown. Using a newly generated CD160 fusion protein (CD160Ig) we examined the role of the novel costimulatory molecule CD160 in mediating CD4(+) or CD8(+) T cell driven allograft rejection. CD160Ig inhibits alloreactive CD8(+) T cell proliferation and IFN-γ production in vitro, in particular in the absence of CD28 costimulation. Consequently CD160Ig prolongs fully mismatched cardiac allograft survival in CD4(-/-), CD28(-/-) knockout and CTLA4Ig treated WT recipients, but not in WT or CD8(-/-) knockout recipients. The prolonged cardiac allograft survival is associated with reduced alloreactive CD8(+) T cell proliferation, effector/memory responses and alloreactive IFN-γ production. Thus, CD160 signaling is particularly important in CD28-independent effector/memory CD8(+) alloreactive T cell activation in vivo and therefore may serve as a novel target for prevention of allograft rejection.
Conflict of interest statement
Figures




Similar articles
-
The impact of CD160 deficiency on alloreactive CD8 T cell responses and allograft rejection.Transl Res. 2022 Jan;239:103-123. doi: 10.1016/j.trsl.2021.08.006. Epub 2021 Aug 27. Transl Res. 2022. PMID: 34461306
-
CD70 signaling is critical for CD28-independent CD8+ T cell-mediated alloimmune responses in vivo.J Immunol. 2005 Feb 1;174(3):1357-64. doi: 10.4049/jimmunol.174.3.1357. J Immunol. 2005. PMID: 15661893
-
Modulation of cytotoxic responses by targeting CD160 prolongs skin graft survival across major histocompatibility class I barrier.Transl Res. 2017 Mar;181:83-95.e3. doi: 10.1016/j.trsl.2016.09.004. Epub 2016 Sep 15. Transl Res. 2017. PMID: 27702550
-
CD160: a unique activating NK cell receptor.Immunol Lett. 2011 Aug 30;138(2):93-6. doi: 10.1016/j.imlet.2011.02.003. Epub 2011 Feb 13. Immunol Lett. 2011. PMID: 21324341 Review.
-
Expression of NK-associated receptors on cytotoxic T cells from melanoma patients: a two-edged sword?Cancer Immunol Immunother. 2004 Oct;53(10):911-24. doi: 10.1007/s00262-004-0507-y. Epub 2004 May 4. Cancer Immunol Immunother. 2004. PMID: 15127235 Free PMC article. Review.
Cited by
-
Increased Heterologous Protein Expression in Drosophila S2 Cells for Massive Production of Immune Ligands/Receptors and Structural Analysis of Human HVEM.Mol Biotechnol. 2015 Oct;57(10):914-22. doi: 10.1007/s12033-015-9881-2. Mol Biotechnol. 2015. PMID: 26202493
-
Identification of CD160-TM as a tumor target on triple negative breast cancers: possible therapeutic applications.Breast Cancer Res. 2024 Feb 15;26(1):28. doi: 10.1186/s13058-024-01785-x. Breast Cancer Res. 2024. PMID: 38360636 Free PMC article.
-
miR-21 antagonism reprograms macrophage metabolism and abrogates chronic allograft vasculopathy.Am J Transplant. 2021 Oct;21(10):3280-3295. doi: 10.1111/ajt.16581. Epub 2021 May 3. Am J Transplant. 2021. PMID: 33764625 Free PMC article.
-
Soluble co-signaling molecules predict long-term graft outcome in kidney-transplanted patients.PLoS One. 2014 Dec 5;9(12):e113396. doi: 10.1371/journal.pone.0113396. eCollection 2014. PLoS One. 2014. PMID: 25478957 Free PMC article.
-
Comprehensively analysis of immunophenotyping signature in triple-negative breast cancer patients based on machine learning.Front Pharmacol. 2023 Jun 23;14:1195864. doi: 10.3389/fphar.2023.1195864. eCollection 2023. Front Pharmacol. 2023. PMID: 37426809 Free PMC article.
References
-
- Rothstein DM, Sayegh MH (2003) T-cell costimulatory pathways in allograft rejection and tolerance. Immunol Rev Dec 196: 85–108. - PubMed
-
- Pearson TC, Alexander DZ, Winn KJ, Linsley PS, Lowry RP, et al. (1994) Transplantation tolerance induced by CTLA4-Ig. Transplantation Jun 27 57(12): 1701–6. - PubMed
-
- Lenschow DJ, Zeng Y, Hathcock KS, Zuckerman LA, Freeman G, et al. (1995) Inhibition of transplant rejection following treatment with anti-B7–2 and anti-B7–1 antibodies. Transplantation 60(10): 1171–8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
