

Mitotic spindle defects and chromosome mis-segregation induced by LDL/cholesterol-implications for Niemann-Pick C1, Alzheimer's disease, and atherosclerosis
- PMID: 23593294
- PMCID: PMC3625184
- DOI: 10.1371/journal.pone.0060718
Mitotic spindle defects and chromosome mis-segregation induced by LDL/cholesterol-implications for Niemann-Pick C1, Alzheimer's disease, and atherosclerosis
Abstract
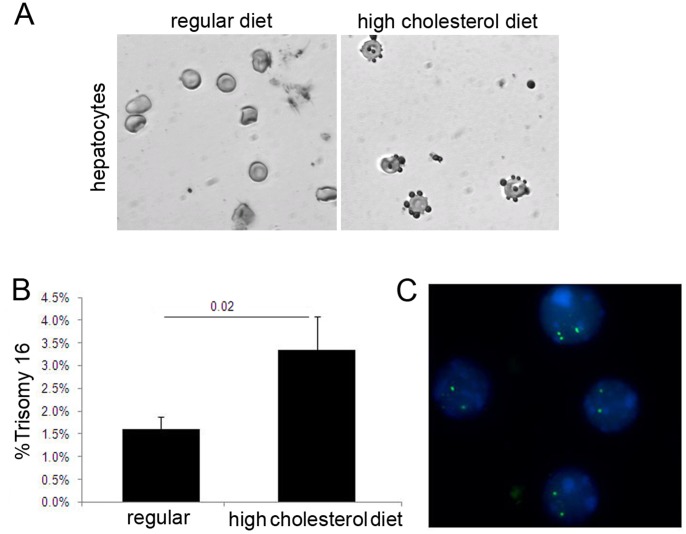
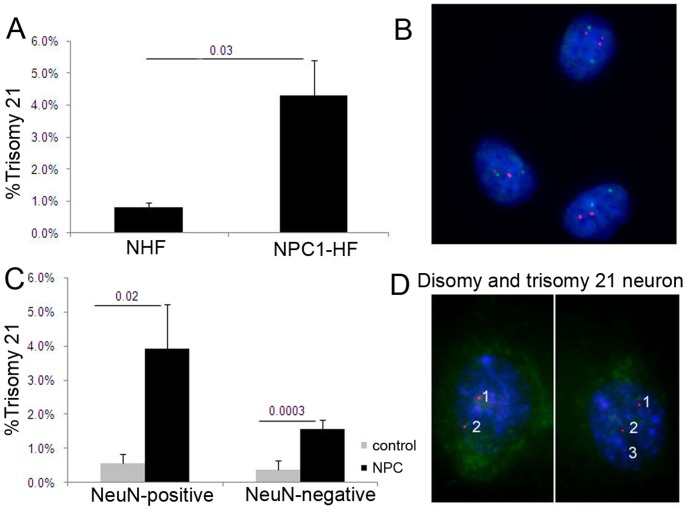
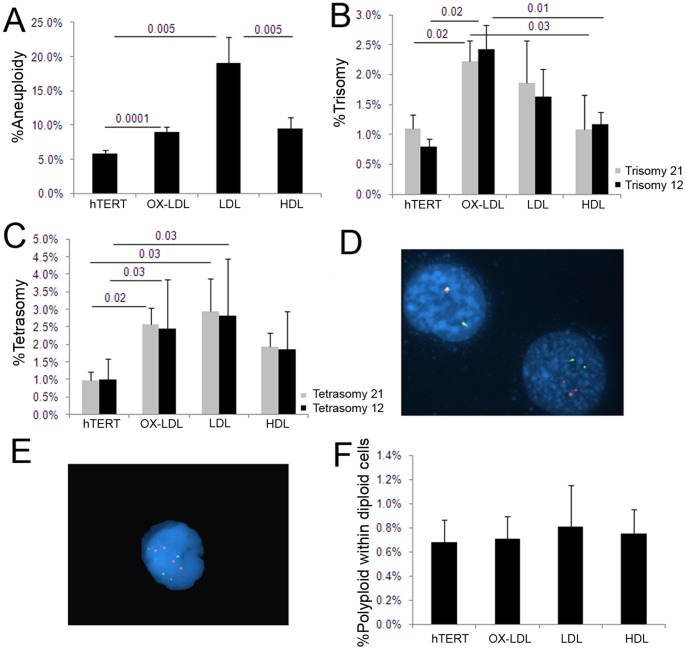
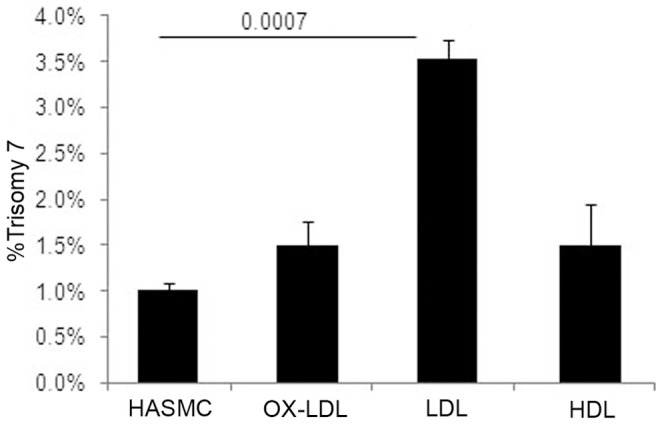
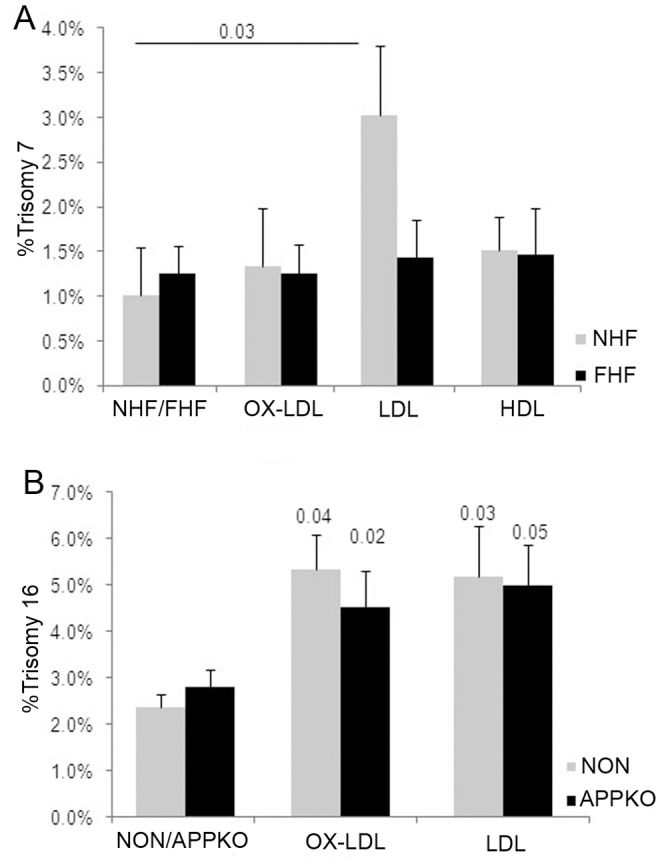
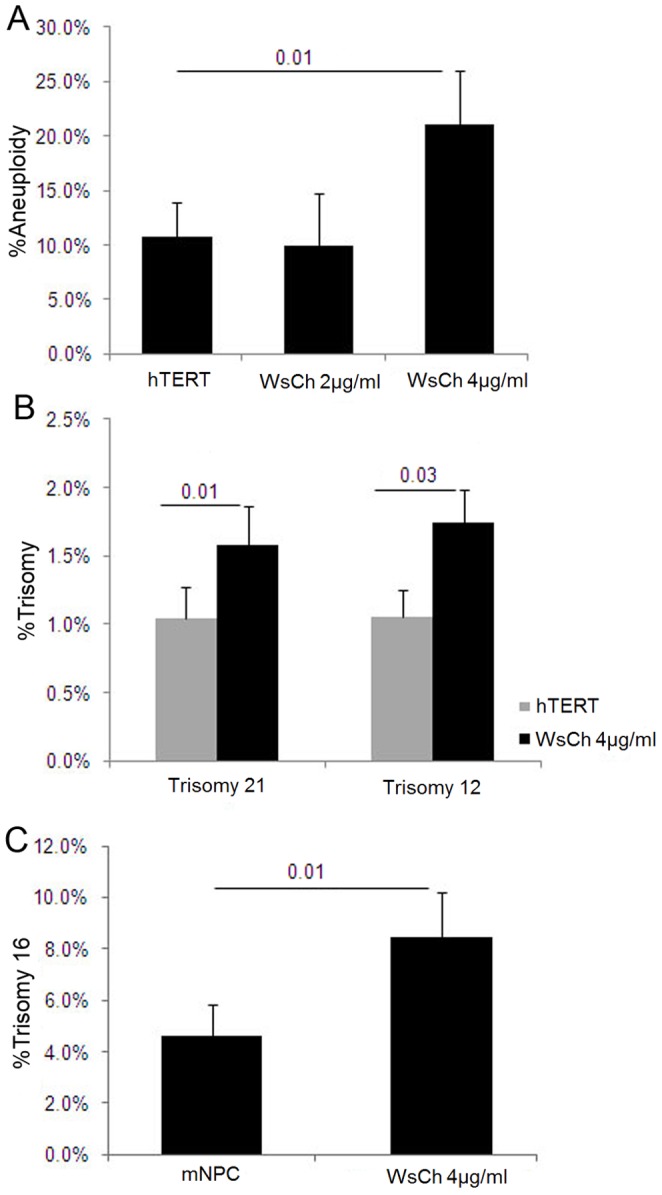
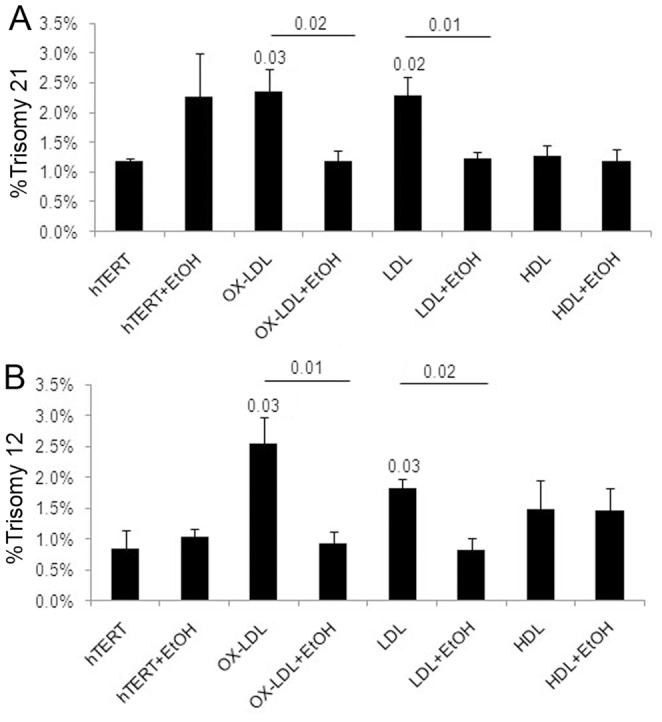
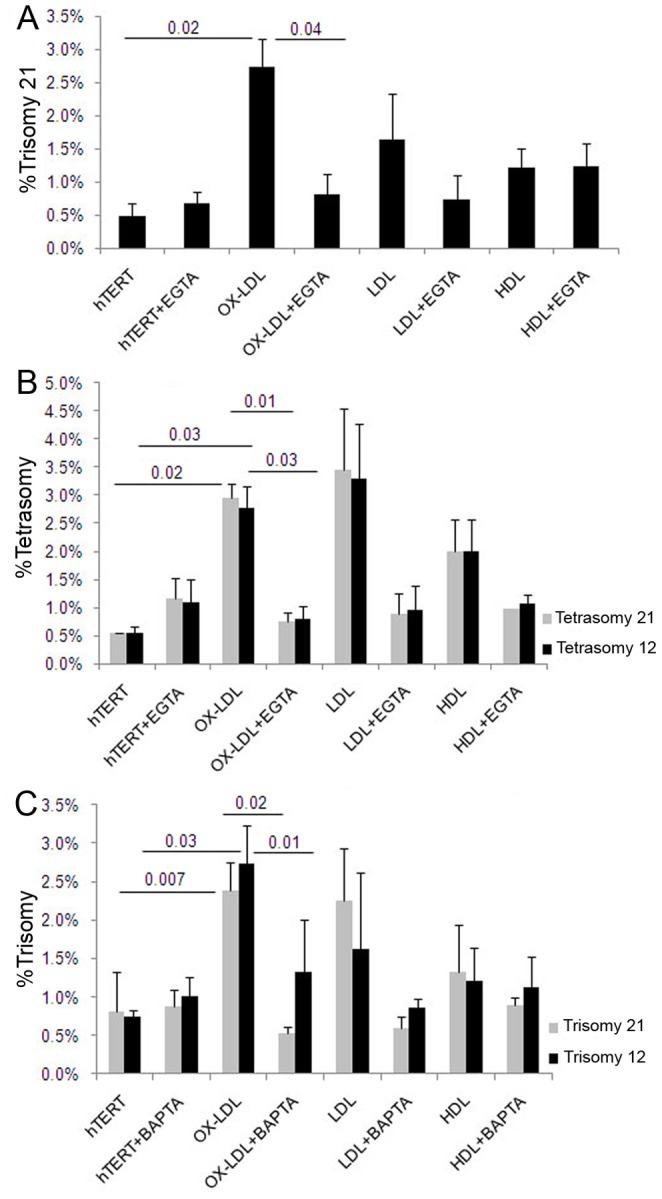
Elevated low-density lipoprotein (LDL)-cholesterol is a risk factor for both Alzheimer's disease (AD) and Atherosclerosis (CVD), suggesting a common lipid-sensitive step in their pathogenesis. Previous results show that AD and CVD also share a cell cycle defect: chromosome instability and up to 30% aneuploidy-in neurons and other cells in AD and in smooth muscle cells in atherosclerotic plaques in CVD. Indeed, specific degeneration of aneuploid neurons accounts for 90% of neuronal loss in AD brain, indicating that aneuploidy underlies AD neurodegeneration. Cell/mouse models of AD develop similar aneuploidy through amyloid-beta (Aß) inhibition of specific microtubule motors and consequent disruption of mitotic spindles. Here we tested the hypothesis that, like upregulated Aß, elevated LDL/cholesterol and altered intracellular cholesterol homeostasis also causes chromosomal instability. Specifically we found that: 1) high dietary cholesterol induces aneuploidy in mice, satisfying the hypothesis' first prediction, 2) Niemann-Pick C1 patients accumulate aneuploid fibroblasts, neurons, and glia, demonstrating a similar aneugenic effect of intracellular cholesterol accumulation in humans 3) oxidized LDL, LDL, and cholesterol, but not high-density lipoprotein (HDL), induce chromosome mis-segregation and aneuploidy in cultured cells, including neuronal precursors, indicating that LDL/cholesterol directly affects the cell cycle, 4) LDL-induced aneuploidy requires the LDL receptor, but not Aß, showing that LDL works differently than Aß, with the same end result, 5) cholesterol treatment disrupts the structure of the mitotic spindle, providing a cell biological mechanism for its aneugenic activity, and 6) ethanol or calcium chelation attenuates lipoprotein-induced chromosome mis-segregation, providing molecular insights into cholesterol's aneugenic mechanism, specifically through its rigidifying effect on the cell membrane, and potentially explaining why ethanol consumption reduces the risk of developing atherosclerosis or AD. These results suggest a novel, cell cycle mechanism by which aberrant cholesterol homeostasis promotes neurodegeneration and atherosclerosis by disrupting chromosome segregation and potentially other aspects of microtubule physiology.
Conflict of interest statement
Figures
Similar articles
-
Alzheimer Abeta peptide induces chromosome mis-segregation and aneuploidy, including trisomy 21: requirement for tau and APP.Mol Biol Cell. 2010 Feb 15;21(4):511-20. doi: 10.1091/mbc.e09-10-0850. Epub 2009 Dec 23. Mol Biol Cell. 2010. PMID: 20032300 Free PMC article.
-
Role of Trisomy 21 Mosaicism in Sporadic and Familial Alzheimer's Disease.Curr Alzheimer Res. 2016;13(1):7-17. doi: 10.2174/156720501301151207100616. Curr Alzheimer Res. 2016. PMID: 26651340 Free PMC article. Review.
-
Alzheimer Aβ disrupts the mitotic spindle and directly inhibits mitotic microtubule motors.Cell Cycle. 2011 May 1;10(9):1397-410. doi: 10.4161/cc.10.9.15478. Epub 2011 May 1. Cell Cycle. 2011. PMID: 21566458 Free PMC article.
-
Alzheimer's presenilin 1 causes chromosome missegregation and aneuploidy.Neurobiol Aging. 2008 Mar;29(3):319-28. doi: 10.1016/j.neurobiolaging.2006.10.027. Epub 2006 Dec 13. Neurobiol Aging. 2008. PMID: 17169464 Free PMC article.
-
Cellular mechanism of U18666A-mediated apoptosis in cultured murine cortical neurons: bridging Niemann-Pick disease type C and Alzheimer's disease.Cell Signal. 2006 Nov;18(11):1844-53. doi: 10.1016/j.cellsig.2006.04.006. Epub 2006 May 7. Cell Signal. 2006. PMID: 16797161 Review.
Cited by
-
Gene and MicroRNA transcriptome analysis of Parkinson's related LRRK2 mouse models.PLoS One. 2014 Jan 10;9(1):e85510. doi: 10.1371/journal.pone.0085510. eCollection 2014. PLoS One. 2014. PMID: 24427314 Free PMC article.
-
Mitotic defects lead to neuronal aneuploidy and apoptosis in frontotemporal lobar degeneration caused by MAPT mutations.Mol Biol Cell. 2018 Mar 1;29(5):575-586. doi: 10.1091/mbc.E17-01-0031. Epub 2017 Dec 27. Mol Biol Cell. 2018. PMID: 29282277 Free PMC article.
-
Study of genotoxic and cytotoxic effects induced in human fibroblasts by exposure to pulsed and continuous 1.6 GHz radiofrequency.Front Public Health. 2024 Jul 31;12:1419525. doi: 10.3389/fpubh.2024.1419525. eCollection 2024. Front Public Health. 2024. PMID: 39145180 Free PMC article.
-
A sporadic Alzheimer's blood-brain barrier model for developing ultrasound-mediated delivery of Aducanumab and anti-Tau antibodies.Theranostics. 2022 Sep 25;12(16):6826-6847. doi: 10.7150/thno.72685. eCollection 2022. Theranostics. 2022. PMID: 36276649 Free PMC article.
-
A Number of the N-terminal RASSF Family: RASSF7.Anticancer Agents Med Chem. 2024;24(12):889-895. doi: 10.2174/1871520622666220930094149. Anticancer Agents Med Chem. 2024. PMID: 36200241
References
-
- Anstey KJ, Lipnicki DM, Low LF (2008) Cholesterol as a risk factor for dementia and cognitive decline: A systematic review of prospective studies with meta-analysis. Am J Geriatr Psychiatry 16: 343–354. - PubMed
-
- Steinberg D (2002) Atherogenesis in perspective: Hypercholesterolemia and inflammation as partners in crime. Nat Med 8: 1211–1217. - PubMed
-
- Querfurth HW, LaFerla HW (2010) Alzheimer’s disease. N Engl J Med 362: 329–344. - PubMed
-
- Hardy J (2006) Alzheimer’s disease: the amyloid cascade hypothesis: an update and reappraisal. J Alzheimers Dis 9: 151–153. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
