

The glutathione reductase GSR-1 determines stress tolerance and longevity in Caenorhabditis elegans
- PMID: 23593298
- PMCID: PMC3620388
- DOI: 10.1371/journal.pone.0060731
The glutathione reductase GSR-1 determines stress tolerance and longevity in Caenorhabditis elegans
Abstract
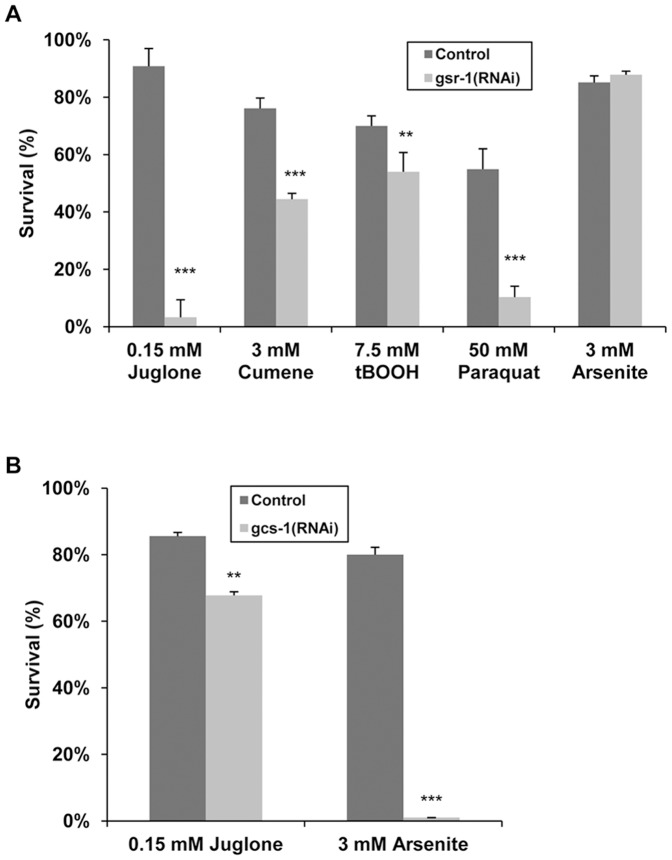
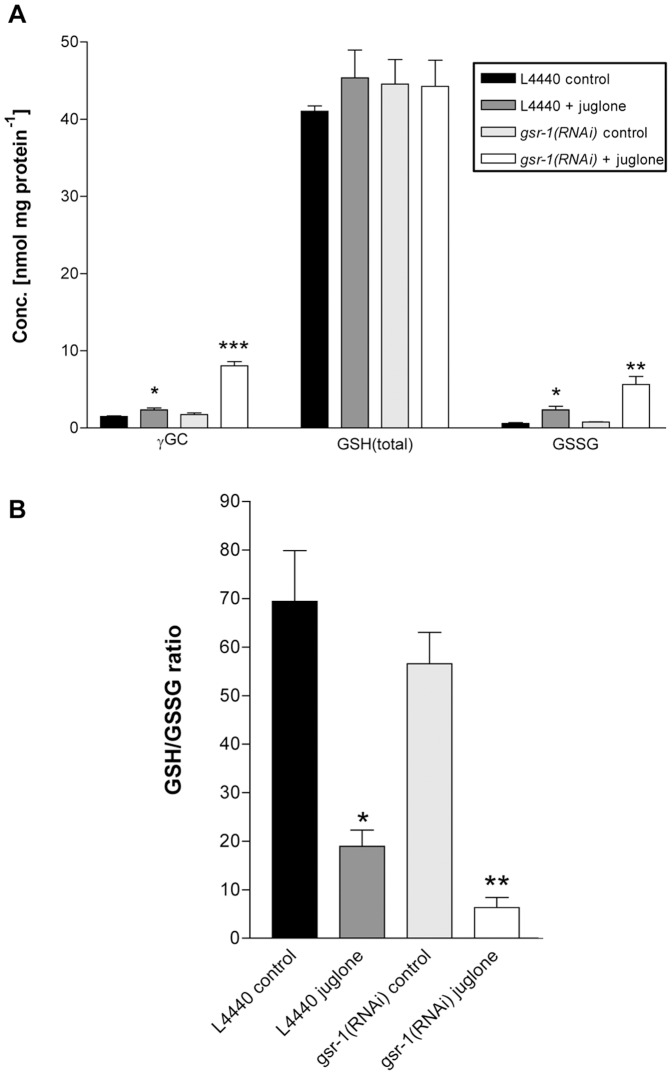
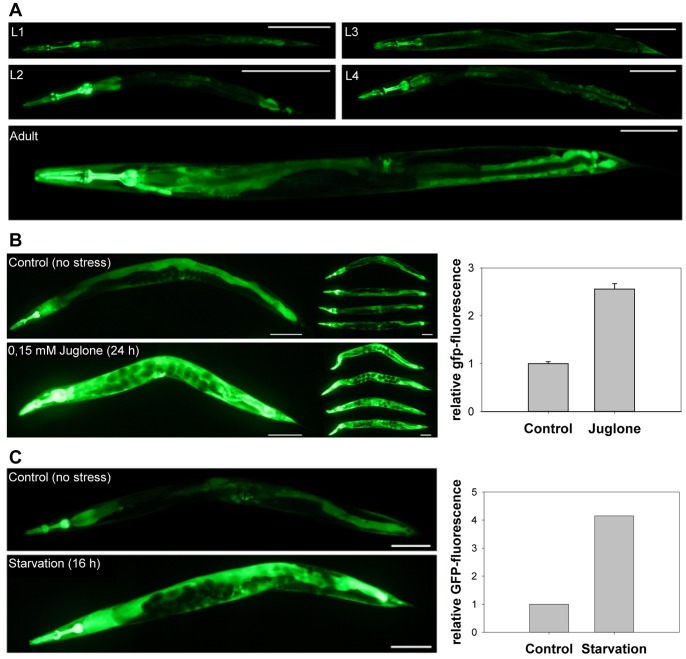
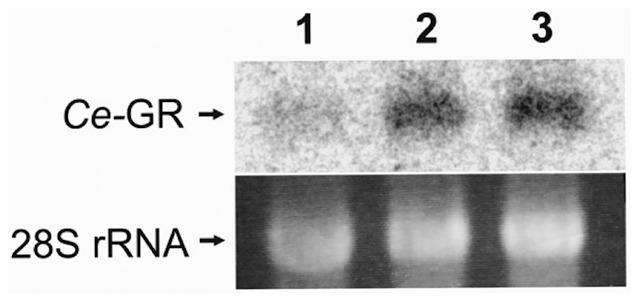
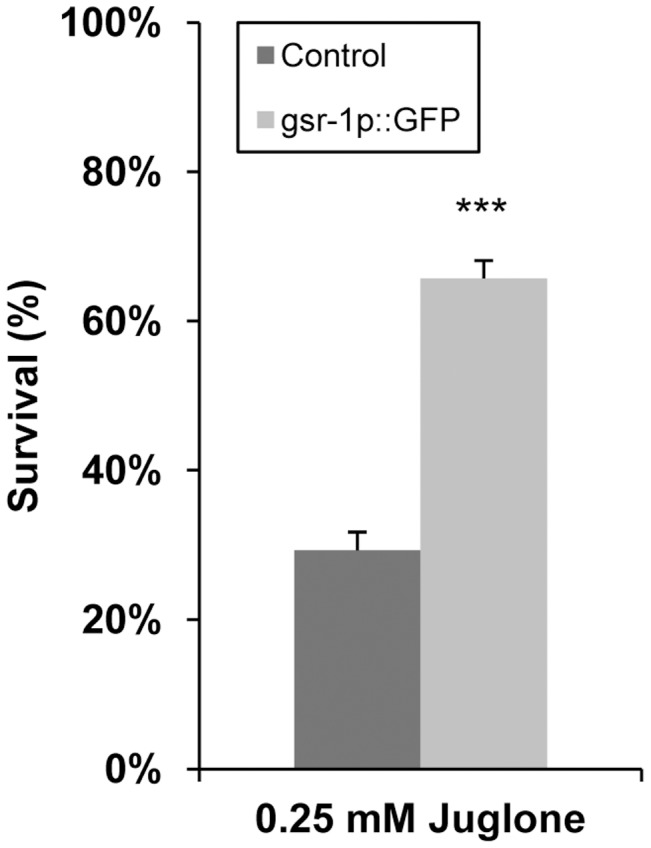
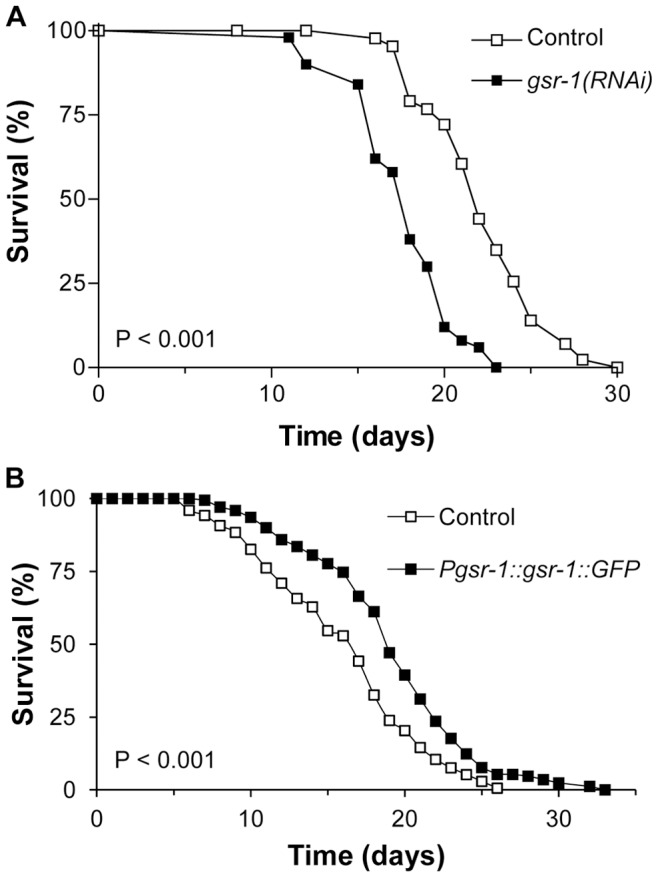
Glutathione (GSH) and GSH-dependent enzymes play a key role in cellular detoxification processes that enable organism to cope with various internal and environmental stressors. However, it is often not clear, which components of the complex GSH-metabolism are required for tolerance towards a certain stressor. To address this question, a small scale RNAi-screen was carried out in Caenorhabditis elegans where GSH-related genes were systematically knocked down and worms were subsequently analysed for their survival rate under sub-lethal concentrations of arsenite and the redox cycler juglone. While the knockdown of γ-glutamylcysteine synthetase led to a diminished survival rate under arsenite stress conditions, GSR-1 (glutathione reductase) was shown to be essential for survival under juglone stress conditions. gsr-1 is the sole GSR encoding gene found in C. elegans. Knockdown of GSR-1 hardly affected total glutathione levels nor reduced glutathione/glutathione disulphide (GSH/GSSG) ratio under normal laboratory conditions. Nevertheless, when GSSG recycling was impaired by gsr-1(RNAi), GSH synthesis was induced, but not vice versa. Moreover, the impact of GSSG recycling was potentiated under oxidative stress conditions, explaining the enormous effect gsr-1(RNAi) knockdown had on juglone tolerance. Accordingly, overexpression of GSR-1 was capable of increasing stress tolerance. Furthermore, expression levels of SKN-1-regulated GSR-1 also affected life span of C. elegans, emphasising the crucial role the GSH redox state plays in both processes.
Conflict of interest statement
Figures
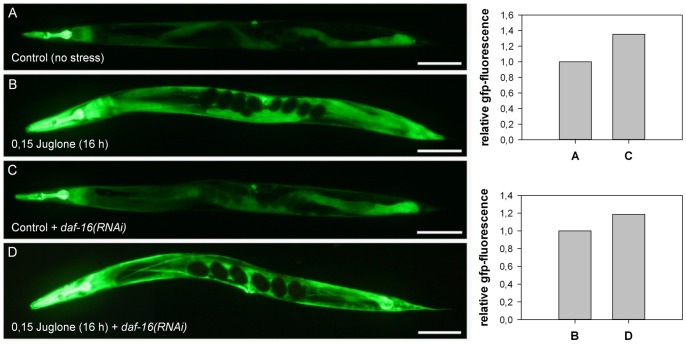
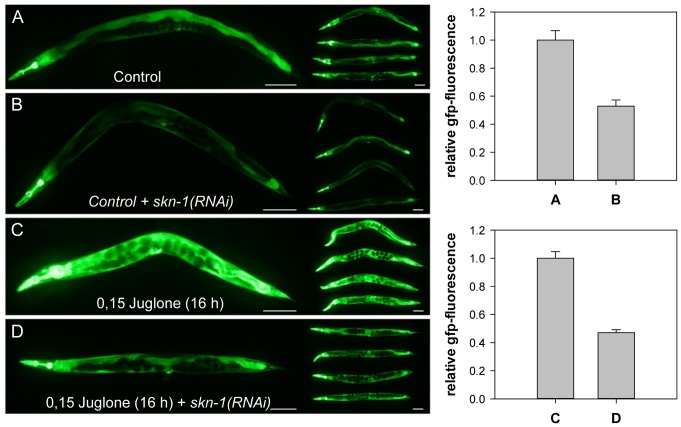
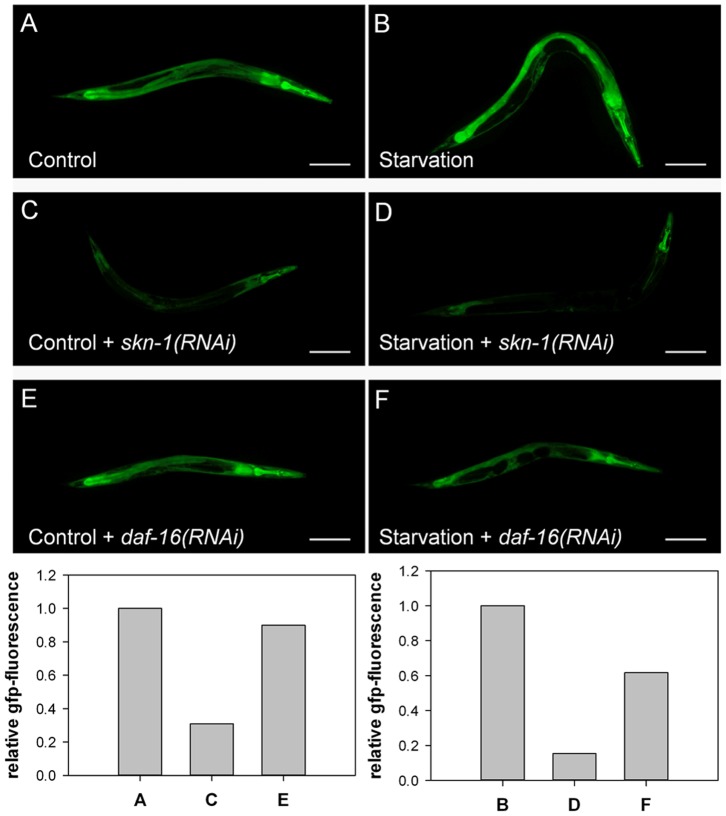
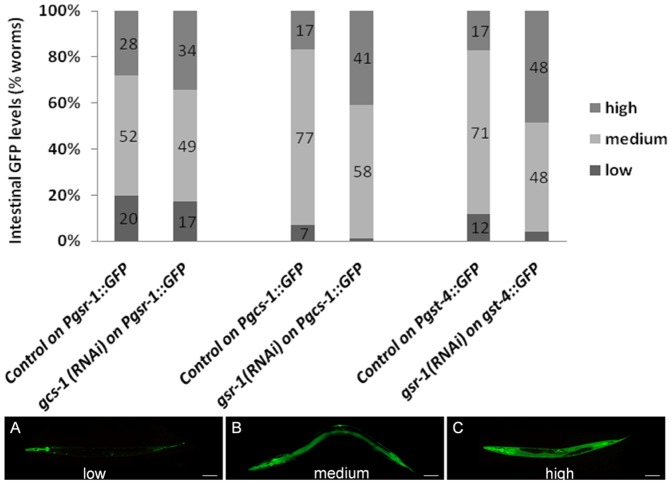
Similar articles
-
The glutathione system and the related thiol network in Caenorhabditis elegans.Redox Biol. 2019 Jun;24:101171. doi: 10.1016/j.redox.2019.101171. Epub 2019 Mar 16. Redox Biol. 2019. PMID: 30901603 Free PMC article. Review.
-
Glutathione reductase gsr-1 is an essential gene required for Caenorhabditis elegans early embryonic development.Free Radic Biol Med. 2016 Jul;96:446-61. doi: 10.1016/j.freeradbiomed.2016.04.017. Epub 2016 Apr 24. Free Radic Biol Med. 2016. PMID: 27117030 Free PMC article.
-
Non-linear impact of glutathione depletion on C. elegans life span and stress resistance.Redox Biol. 2017 Apr;11:502-515. doi: 10.1016/j.redox.2016.12.003. Epub 2016 Dec 6. Redox Biol. 2017. PMID: 28086197 Free PMC article.
-
Evaluation of the antioxidant effects of acid hydrolysates from Auricularia auricular polysaccharides using a Caenorhabditis elegans model.Food Funct. 2019 Sep 1;10(9):5531-5543. doi: 10.1039/c8fo02589d. Epub 2019 Aug 16. Food Funct. 2019. PMID: 31418439
-
Formation and regulation of adaptive response in nematode Caenorhabditis elegans.Oxid Med Cell Longev. 2012;2012:564093. doi: 10.1155/2012/564093. Epub 2012 Sep 11. Oxid Med Cell Longev. 2012. PMID: 22997543 Free PMC article. Review.
Cited by
-
The Secretome of Human Neonatal Mesenchymal Stem Cells Modulates Doxorubicin-Induced Cytotoxicity: Impact in Non-Tumor Cells.Int J Mol Sci. 2021 Dec 3;22(23):13072. doi: 10.3390/ijms222313072. Int J Mol Sci. 2021. PMID: 34884877 Free PMC article.
-
The glutathione system and the related thiol network in Caenorhabditis elegans.Redox Biol. 2019 Jun;24:101171. doi: 10.1016/j.redox.2019.101171. Epub 2019 Mar 16. Redox Biol. 2019. PMID: 30901603 Free PMC article. Review.
-
Genetic diversity and adaptive evolutionary history of Sapindus in China: insights from whole-genome resequencing of 100 representative individuals.Plant Biotechnol J. 2025 Jul;23(7):2485-2500. doi: 10.1111/pbi.70058. Epub 2025 Mar 30. Plant Biotechnol J. 2025. PMID: 40159695 Free PMC article.
-
Glutathione reductase gsr-1 is an essential gene required for Caenorhabditis elegans early embryonic development.Free Radic Biol Med. 2016 Jul;96:446-61. doi: 10.1016/j.freeradbiomed.2016.04.017. Epub 2016 Apr 24. Free Radic Biol Med. 2016. PMID: 27117030 Free PMC article.
-
Biological stability of DNA methylation measurements over varying intervals of time and in the presence of acute stress.Epigenetics. 2023 Dec;18(1):2230686. doi: 10.1080/15592294.2023.2230686. Epigenetics. 2023. PMID: 37393564 Free PMC article.
References
-
- Sies H (1993) Strategies of antioxidant defense. Eur J Biochem 215: 213–219. - PubMed
-
- Davies KJ (2000) Oxidative stress, antioxidant defenses, and damage removal, repair, and replacement systems. IUBMB Life 50: 279–289. - PubMed
-
- Valko M, Leibfritz D, Moncol J, Cronin MT, Mazur M, et al. (2007) Free radicals and antioxidants in normal physiological functions and human disease. Int J Biochem Cell Biol 39: 44–84. - PubMed
-
- Limon-Pacheco J, Gonsebatt ME (2009) The role of antioxidants and antioxidant-related enzymes in protective responses to environmentally induced oxidative stress. Mutat Res 674: 137–147. - PubMed
-
- Harman D (1956) Aging: a theory based on free radical and radiation chemistry. J Gerontol 11: 298–300. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
