

Cortical-like receptive fields in the lateral geniculate nucleus of marmoset monkeys
- PMID: 23595745
- PMCID: PMC6618877
- DOI: 10.1523/JNEUROSCI.5208-12.2013
Cortical-like receptive fields in the lateral geniculate nucleus of marmoset monkeys
Abstract
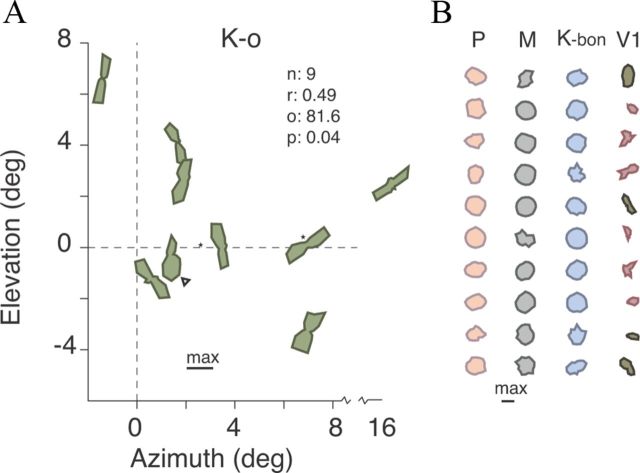
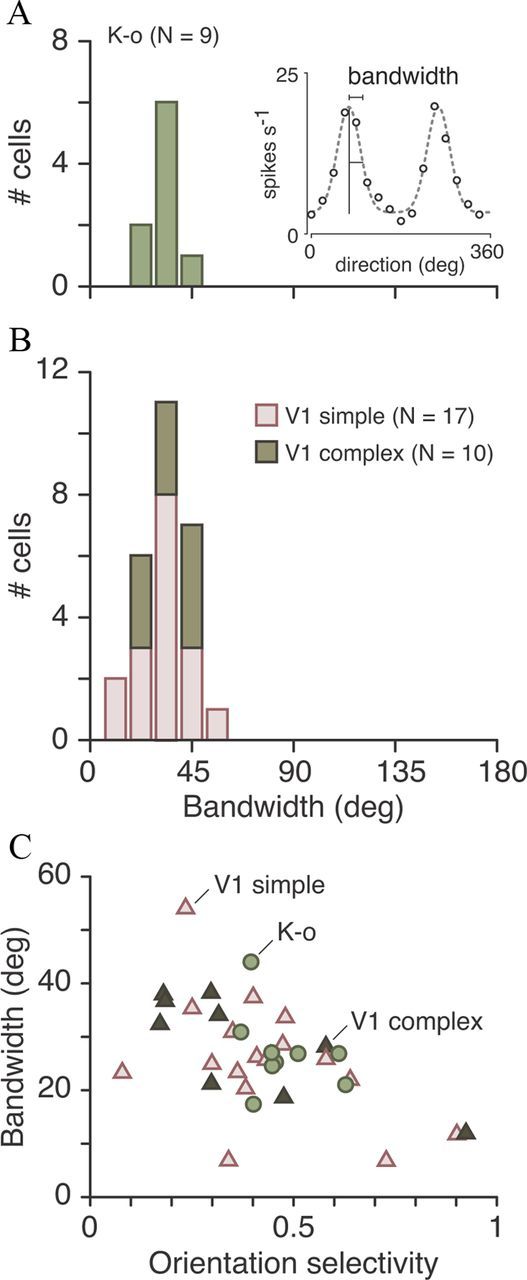
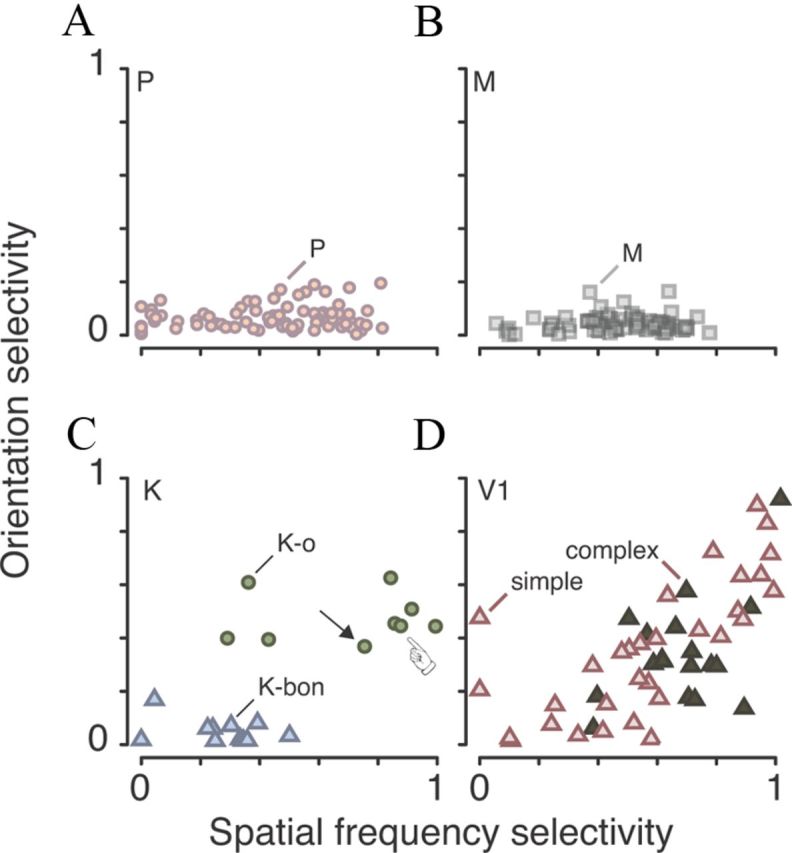
Most neurons in primary visual cortex (V1) exhibit high selectivity for the orientation of visual stimuli. In contrast, neurons in the main thalamic input to V1, the lateral geniculate nucleus (LGN), are considered to be only weakly orientation selective. Here we characterize a sparse population of cells in marmoset LGN that show orientation and spatial frequency selectivity as great as that of cells in V1. The recording position in LGN and histological reconstruction of these cells shows that they are part of the koniocellular (K) pathways. Accordingly we have named them K-o ("koniocellular-orientation") cells. Most K-o cells prefer vertically oriented gratings; their contrast sensitivity and TF tuning are similar to those of parvocellular cells, and they receive negligible functional input from short wavelength-sensitive ("blue") cone photoreceptors. Four K-o cells tested displayed binocular responses. Our results provide further evidence that in primates as in nonprimate mammals the cortical input streams include a diversity of visual representations. The presence of K-o cells increases functional homologies between K pathways in primates and "sluggish/W" pathways in nonprimate visual systems.
Figures










References
-
- Berens P. CircStat: a MATLAB toolbox for circular statistics. J Stat Software. 2009;31
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous