

The kindlin 3 pleckstrin homology domain has an essential role in lymphocyte function-associated antigen 1 (LFA-1) integrin-mediated B cell adhesion and migration
- PMID: 23595985
- PMCID: PMC3663508
- DOI: 10.1074/jbc.M112.434621
The kindlin 3 pleckstrin homology domain has an essential role in lymphocyte function-associated antigen 1 (LFA-1) integrin-mediated B cell adhesion and migration
Abstract
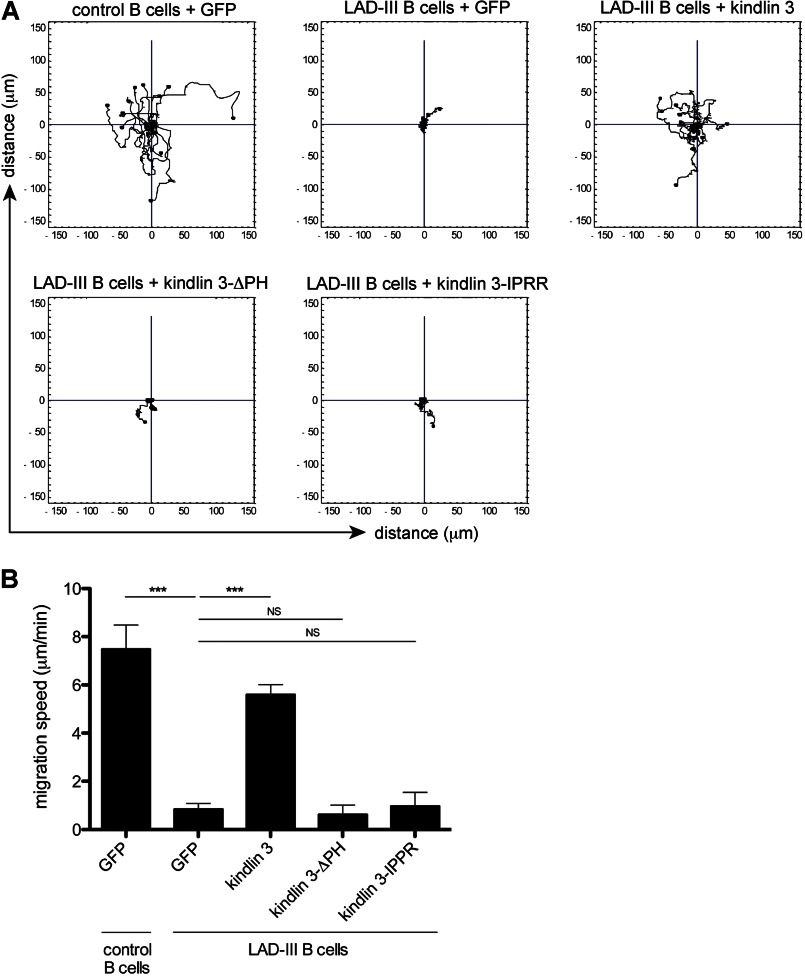
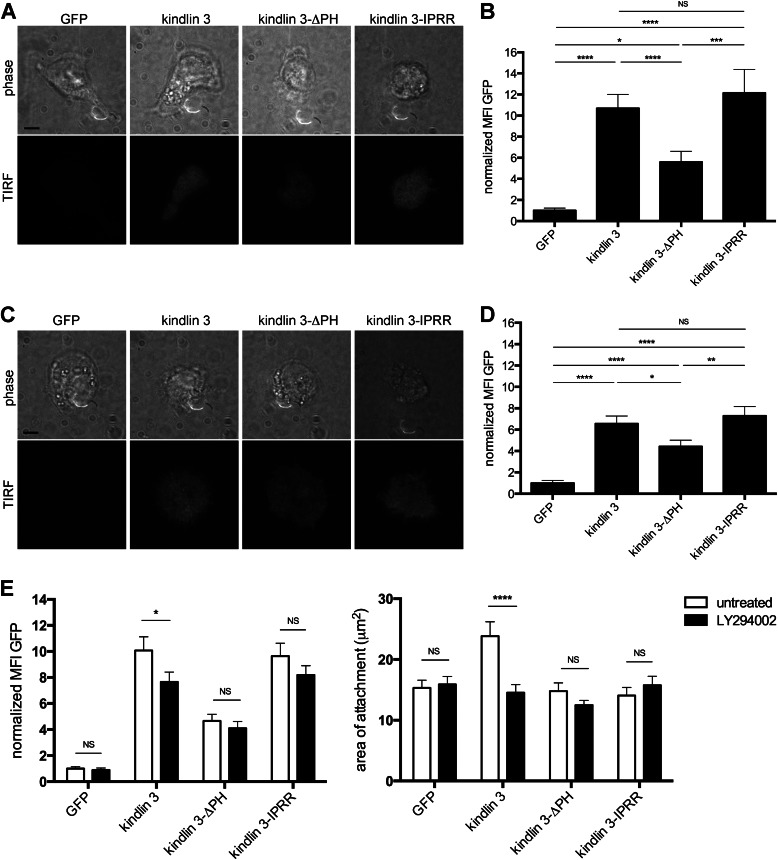
The protein kindlin 3 is mutated in the leukocyte adhesion deficiency III (LAD-III) disorder, leading to widespread infection due to the failure of leukocytes to migrate into infected tissue sites. To gain understanding of how kindlin 3 controls leukocyte function, we have focused on its pleckstrin homology (PH) domain and find that deletion of this domain eliminates the ability of kindlin 3 to participate in adhesion and migration of B cells mediated by the leukocyte integrin lymphocyte function-associated antigen 1 (LFA-1). PH domains are often involved in membrane localization of proteins through binding to phosphoinositides. We show that the kindlin 3 PH domain has binding affinity for phosphoinositide PI(3,4,5)P3 over PI(4,5)P2. It has a major role in membrane association of kindlin 3 that is enhanced by the binding of LFA-1 to intercellular adhesion molecule 1 (ICAM-1). A splice variant, kindlin 3-IPRR, has a four-residue insert in the PH domain at a critical site that influences phosphoinositide binding by enhancing binding to PI(4,5)P2 as well as by binding to PI(3,4,5)P3. However kindlin 3-IPRR is unable to restore the ability of LAD-III B cells to adhere to and migrate on LFA-1 ligand ICAM-1, potentially by altering the dynamics or PI specificity of binding to the membrane. Thus, the correct functioning of the kindlin 3 PH domain is central to the role that kindlin 3 performs in guiding lymphocyte adhesion and motility behavior, which in turn is required for a successful immune response.
Keywords: Adhesion; Cell Migration; Cell Signaling; Integrin; Kindlin; Lymphocyte; PH Domain; Receptors.
Figures



Similar articles
-
Kindlin-3 is required for the stabilization of TCR-stimulated LFA-1:ICAM-1 bonds critical for lymphocyte arrest and spreading on dendritic cells.Blood. 2011 Jun 30;117(26):7042-52. doi: 10.1182/blood-2010-12-322859. Epub 2011 May 2. Blood. 2011. PMID: 21536861
-
Loss of Kindlin-3 in LAD-III eliminates LFA-1 but not VLA-4 adhesiveness developed under shear flow conditions.Blood. 2009 Sep 10;114(11):2344-53. doi: 10.1182/blood-2009-04-218636. Epub 2009 Jul 17. Blood. 2009. PMID: 19617577
-
Examining the Effect of Kindlin-3 Binding Site Mutation on LFA-1-ICAM-1 Bonds by Force Measuring Optical Tweezers.Front Immunol. 2022 Jan 26;12:792813. doi: 10.3389/fimmu.2021.792813. eCollection 2021. Front Immunol. 2022. PMID: 35154074 Free PMC article.
-
Inhibitors targeting the LFA-1/ICAM-1 cell-adhesion interaction: design and mechanism of action.Curr Pharm Des. 2008;14(22):2128-39. doi: 10.2174/138161208785740225. Curr Pharm Des. 2008. PMID: 18781967 Review.
-
Paxillin and Kindlin: Research Progress and Biological Functions.Biomolecules. 2025 Jan 24;15(2):173. doi: 10.3390/biom15020173. Biomolecules. 2025. PMID: 40001476 Free PMC article. Review.
Cited by
-
NMR Characterization and Membrane Interactions of the Loop Region of Kindlin-3 F1 Subdomain.PLoS One. 2016 Apr 21;11(4):e0153501. doi: 10.1371/journal.pone.0153501. eCollection 2016. PLoS One. 2016. PMID: 27101375 Free PMC article.
-
Talin and kindlin: the one-two punch in integrin activation.Front Med. 2014 Mar;8(1):6-16. doi: 10.1007/s11684-014-0317-3. Epub 2014 Jan 29. Front Med. 2014. PMID: 24477625 Review.
-
Paxillin binding to the PH domain of kindlin-3 in platelets is required to support integrin αIIbβ3 outside-in signaling.J Thromb Haemost. 2021 Dec;19(12):3126-3138. doi: 10.1111/jth.15505. Epub 2021 Aug 31. J Thromb Haemost. 2021. PMID: 34411430 Free PMC article.
-
Kindlin-3 recruitment to the plasma membrane precedes high-affinity β2-integrin and neutrophil arrest from rolling.Blood. 2021 Jan 7;137(1):29-38. doi: 10.1182/blood.2019003446. Blood. 2021. PMID: 32777822 Free PMC article.
-
The Interplay between Integrins and Immune Cells as a Regulator in Cancer Immunology.Int J Mol Sci. 2023 Mar 24;24(7):6170. doi: 10.3390/ijms24076170. Int J Mol Sci. 2023. PMID: 37047140 Free PMC article. Review.
References
-
- Kinashi T. (2005) Intracellular signalling controlling integrin activation in lymphocytes. Nat. Rev. Immunol. 5, 546–559 - PubMed
-
- Alon R., Dustin M. L. (2007) Force as a facilitator of integrin conformational changes during leukocyte arrest on blood vessels and antigen-presenting cells. Immunity 26, 17–27 - PubMed
-
- Hogg N., Patzak I., Willenbrock F. (2011) The insider's guide to leukocyte integrin signalling and function. Nat. Rev. Immunol. 11, 416–426 - PubMed
-
- Etzioni A., Alon R. (2004) Leukocyte adhesion deficiency III: a group of integrin activation defects in hematopoietic lineage cells. Curr. Opin. Allergy Clin. Immunol. 4, 485–490 - PubMed
-
- van de Vijver E., Maddalena A., Sanal Ö., Holland S. M., Uzel G., Madkaikar M., de Boer M., van Leeuwen K., Köker M. Y., Parvaneh N., Fischer A., Law S. K., Klein N., Tezcan F. I., Unal E., Patiroglu T., Belohradsky B. H., Schwartz K., Somech R., Kuijpers T. W., Roos D. (2012) Hematologically important mutations: leukocyte adhesion deficiency (first update). Blood Cells Mol. Dis. 48, 53–61 - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
