

Molecular view of an electron transfer process essential for iron-sulfur protein biogenesis
- PMID: 23596212
- PMCID: PMC3645582
- DOI: 10.1073/pnas.1302378110
Molecular view of an electron transfer process essential for iron-sulfur protein biogenesis
Abstract
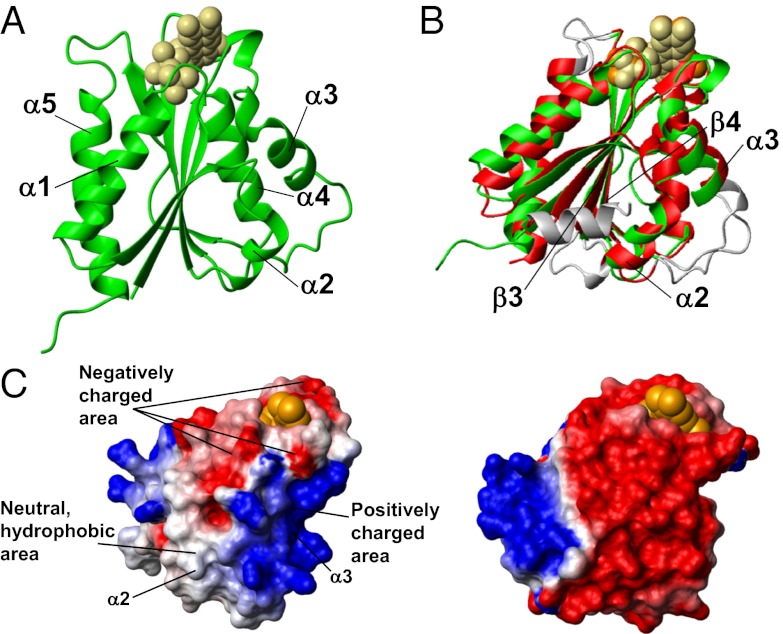
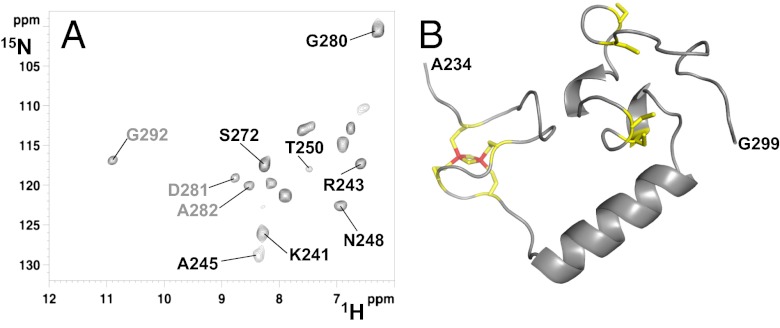
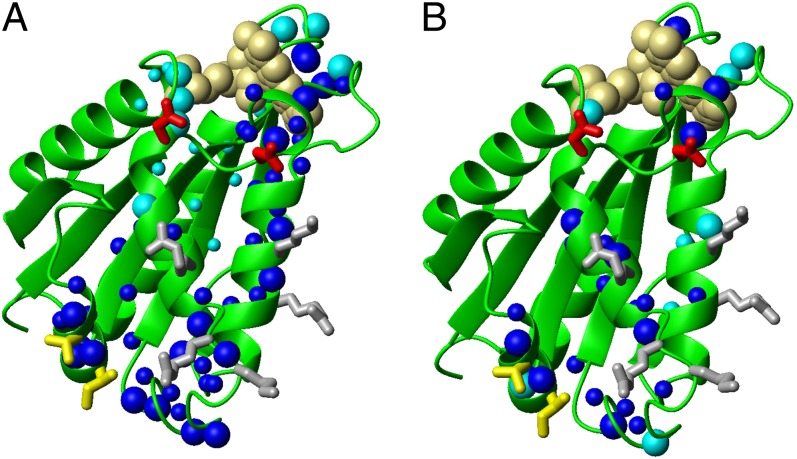
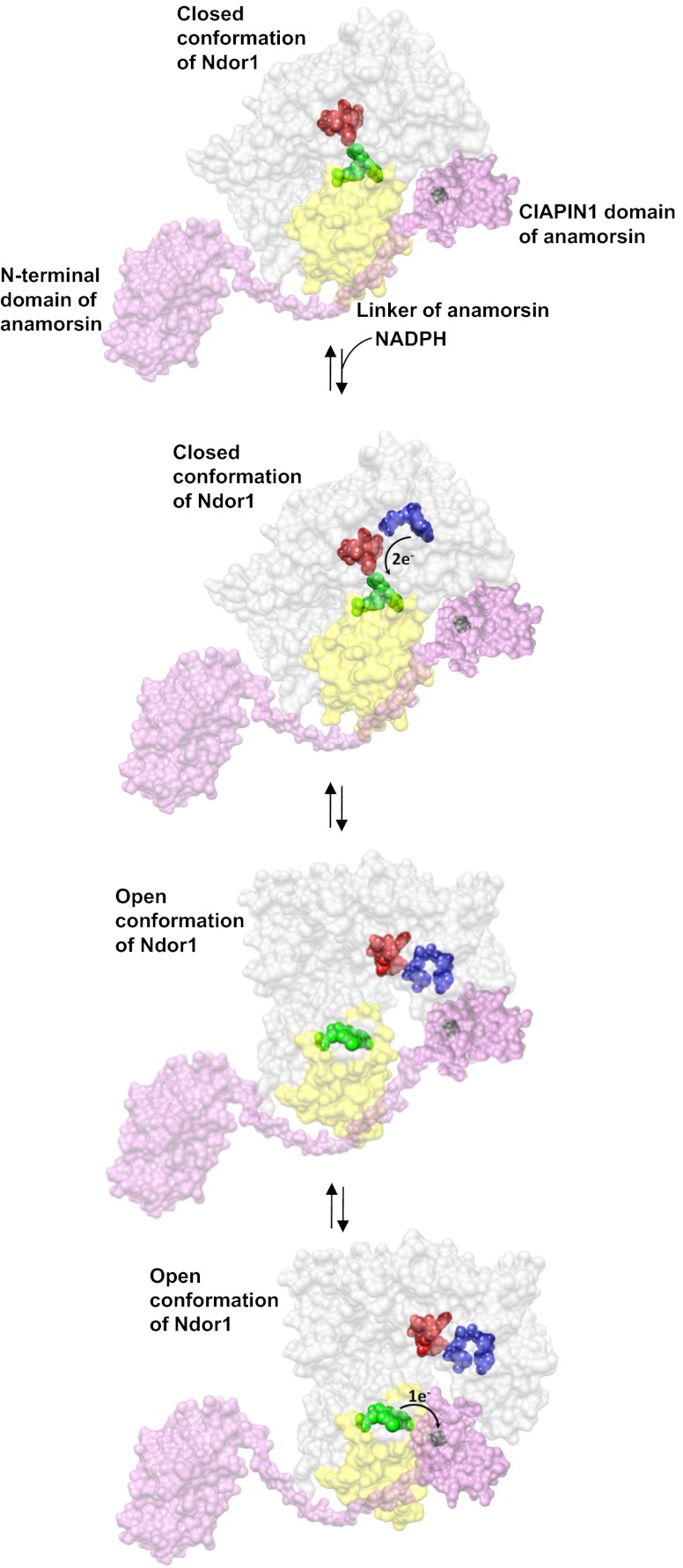
Biogenesis of iron-sulfur cluster proteins is a highly regulated process that requires complex protein machineries. In the cytosolic iron-sulfur protein assembly machinery, two human key proteins--NADPH-dependent diflavin oxidoreductase 1 (Ndor1) and anamorsin--form a stable complex in vivo that was proposed to provide electrons for assembling cytosolic iron-sulfur cluster proteins. The Ndor1-anamorsin interaction was also suggested to be implicated in the regulation of cell survival/death mechanisms. In the present work we unravel the molecular basis of recognition between Ndor1 and anamorsin and of the electron transfer process. This is based on the structural characterization of the two partner proteins, the investigation of the electron transfer process, and the identification of those protein regions involved in complex formation and those involved in electron transfer. We found that an unstructured region of anamorsin is essential for the formation of a specific and stable protein complex with Ndor1, whereas the C-terminal region of anamorsin, containing the [2Fe-2S] redox center, transiently interacts through complementary charged residues with the FMN-binding site region of Ndor1 to perform electron transfer. Our results propose a molecular model of the electron transfer process that is crucial for understanding the functional role of this interaction in human cells.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Johnson DC, Dean DR, Smith AD, Johnson MK. Structure, function, and formation of biological iron-sulfur clusters. Annu Rev Biochem. 2005;74:247–281. - PubMed
-
- Lill R. Function and biogenesis of iron-sulphur proteins. Nature. 2009;460(7257):831–838. - PubMed
-
- Meyer J. Iron-sulfur protein folds, iron-sulfur chemistry, and evolution. J Biol Inorg Chem. 2008;13(2):157–170. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
