

Trafficking of the Menkes copper transporter ATP7A is regulated by clathrin-, AP-2-, AP-1-, and Rab22-dependent steps
- PMID: 23596324
- PMCID: PMC3667726
- DOI: 10.1091/mbc.E12-08-0625
Trafficking of the Menkes copper transporter ATP7A is regulated by clathrin-, AP-2-, AP-1-, and Rab22-dependent steps
Abstract
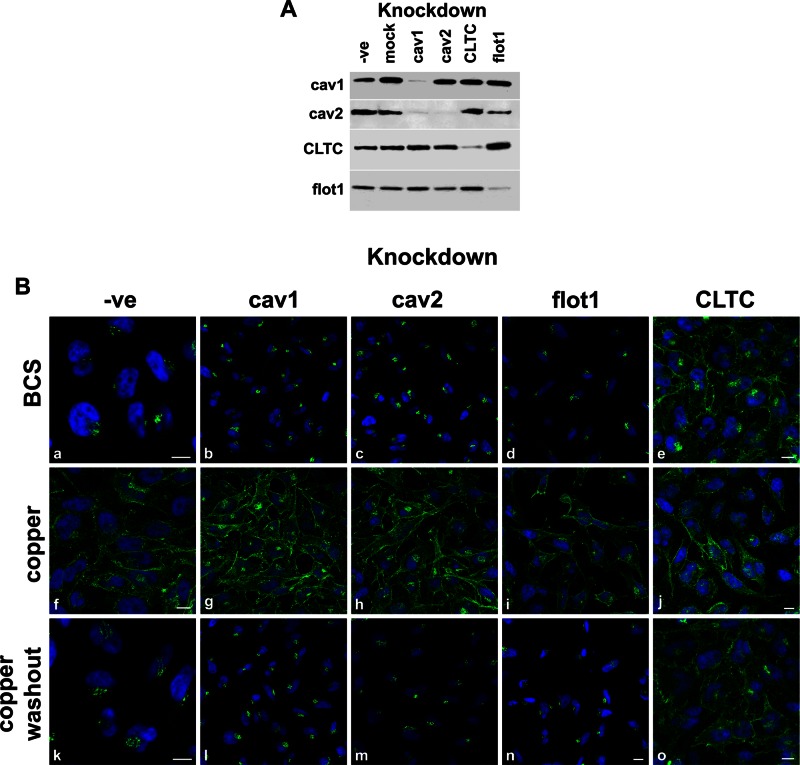
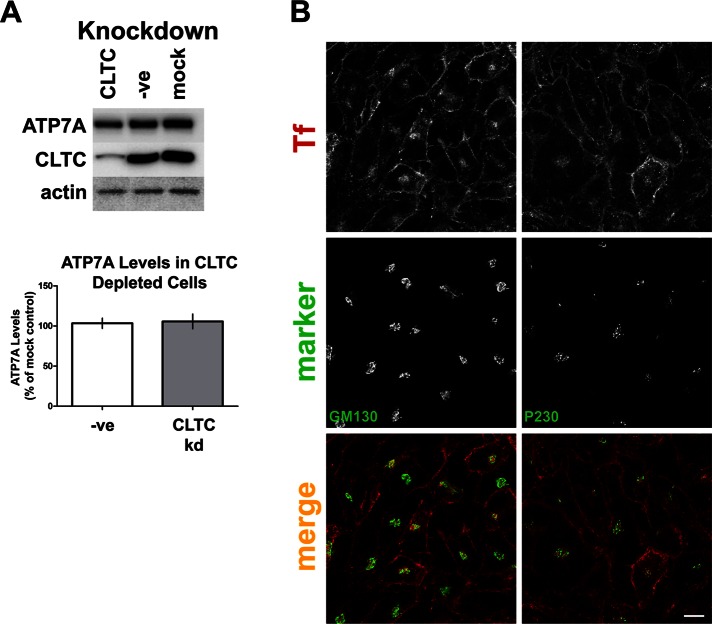
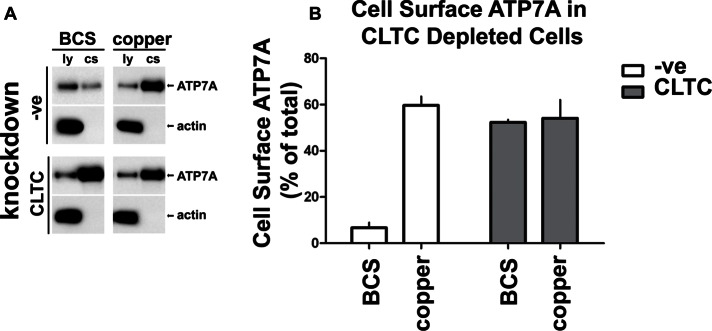
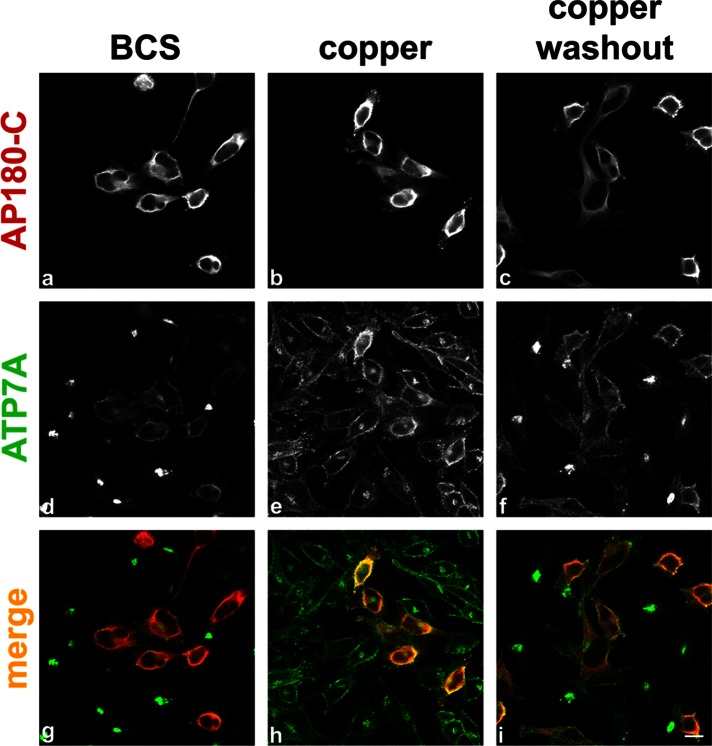
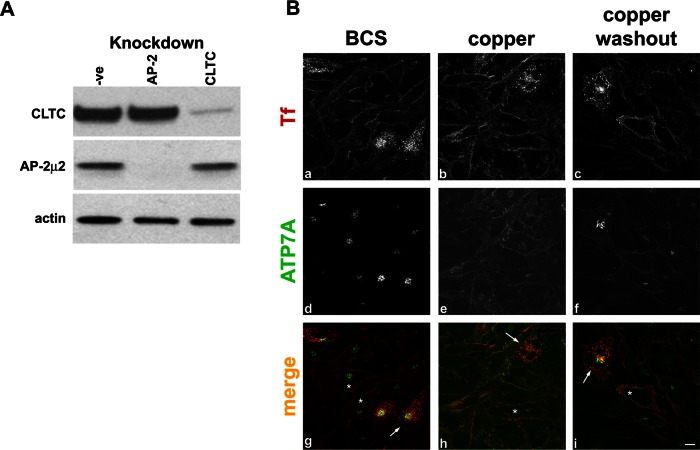
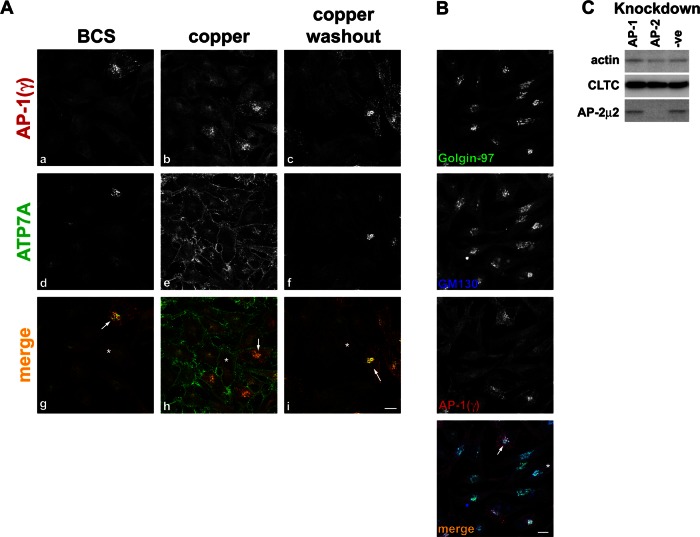
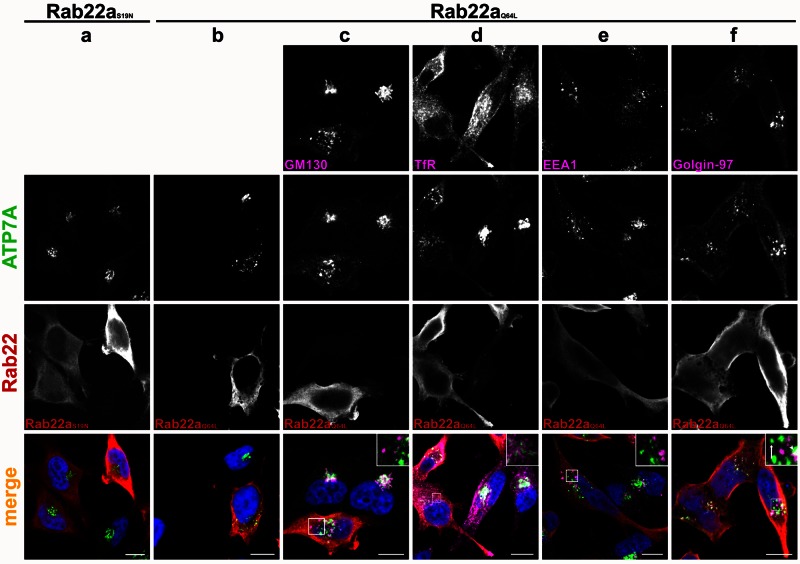
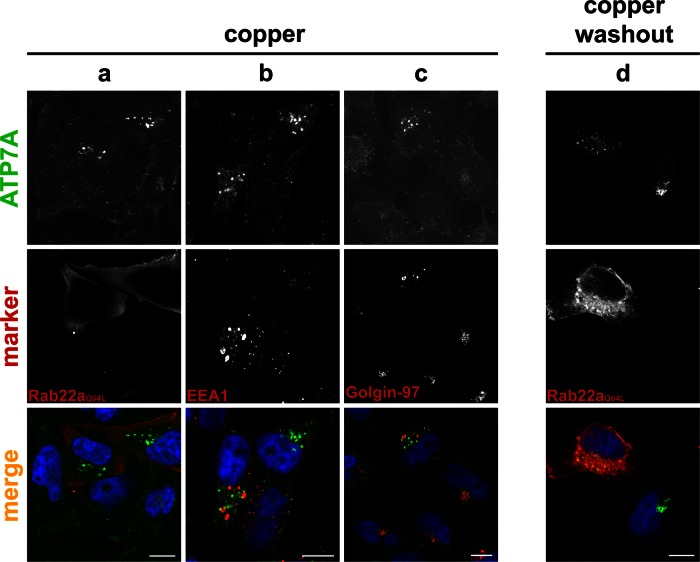
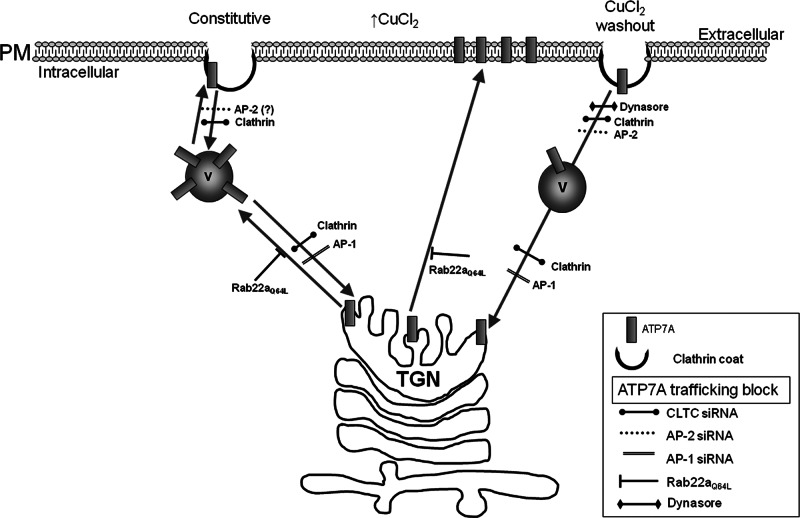
The transporter ATP7A mediates systemic copper absorption and provides cuproenzymes in the trans-Golgi network (TGN) with copper. To regulate metal homeostasis, ATP7A constitutively cycles between the TGN and plasma membrane (PM). ATP7A trafficking to the PM is elevated in response to increased copper load and is reversed when copper concentrations are lowered. Molecular mechanisms underlying this trafficking are poorly understood. We assess the role of clathrin, adaptor complexes, lipid rafts, and Rab22a in an attempt to decipher the regulatory proteins involved in ATP7A cycling. While RNA interference (RNAi)-mediated depletion of caveolin 1/2 or flotillin had no effect on ATP7A localization, clathrin heavy chain depletion or expression of AP180 dominant-negative mutant not only disrupted clathrin-regulated pathways, but also blocked PM-to-TGN internalization of ATP7A. Depletion of the μ subunits of either adaptor protein-2 (AP-2) or AP-1 using RNAi further provides evidence that both clathrin adaptors are important for trafficking of ATP7A from the PM to the TGN. Expression of the GTP-locked Rab22aQ64L mutant caused fragmentation of TGN membrane domains enriched for ATP7A. These appear to be a subdomain of the mammalian TGN, showing only partial overlap with the TGN marker golgin-97. Of importance, ATP7A remained in the Rab22aQ64L-generated structures after copper treatment and washout, suggesting that forward trafficking out of this compartment was blocked. This study provides evidence that multiple membrane-associated factors, including clathrin, AP-2, AP-1, and Rab22, are regulators of ATP7A trafficking.
Figures
References
-
- Balamurugan K, Schaffner W. Copper homeostasis in eukaryotes: teetering on a tightrope. Biochim Biophys Acta. 2006;1763:737–746. - PubMed
-
- Bhatnagar A, Sheffler DJ, Kroeze WK, Compton-Toth B, Roth BL. Caveolin-1 interacts with 5-HT2A serotonin receptors and profoundly modulates the signaling of selected Galphaq-coupled protein receptors. J Biol Chem. 2004;279:34614–34623. - PubMed
-
- Bonifacino JS, Traub LM. Signals for sorting of transmembrane proteins to endosomes and lysosomes. Annu Rev Biochem. 2003;72:395–447. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
