

Cortical speech-evoked response patterns in multiple auditory fields are correlated with behavioral discrimination ability
- PMID: 23596332
- PMCID: PMC3727033
- DOI: 10.1152/jn.00092.2013
Cortical speech-evoked response patterns in multiple auditory fields are correlated with behavioral discrimination ability
Abstract
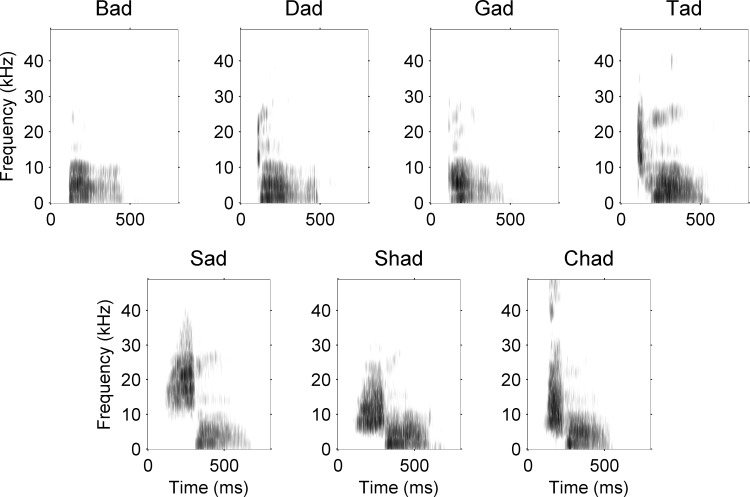
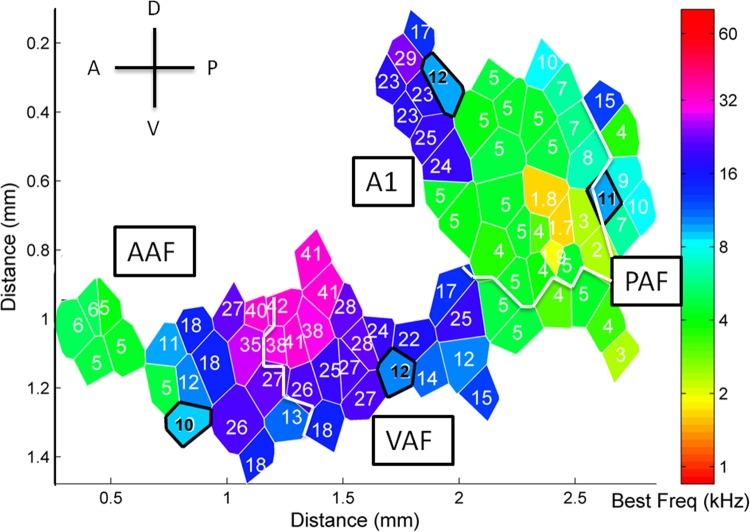
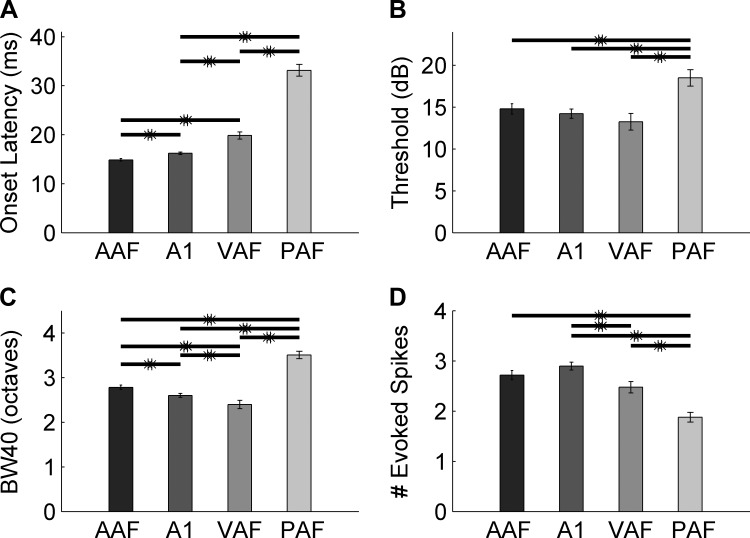
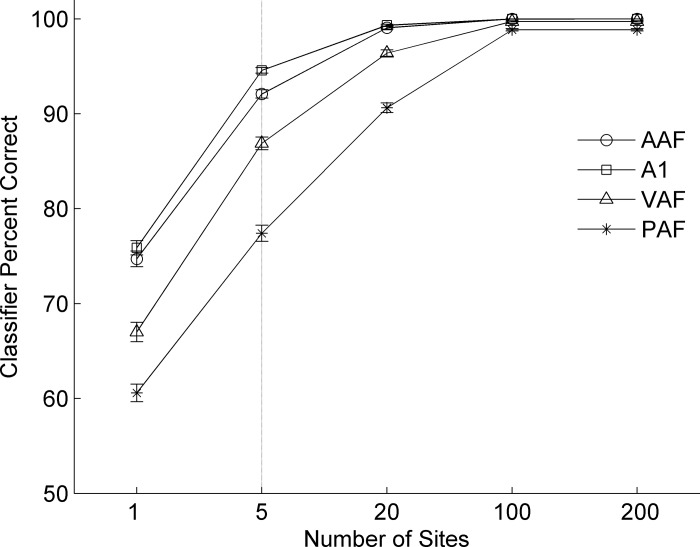
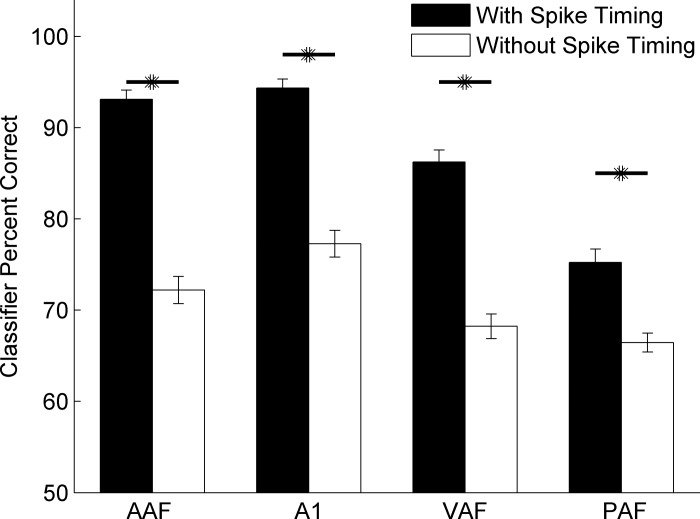
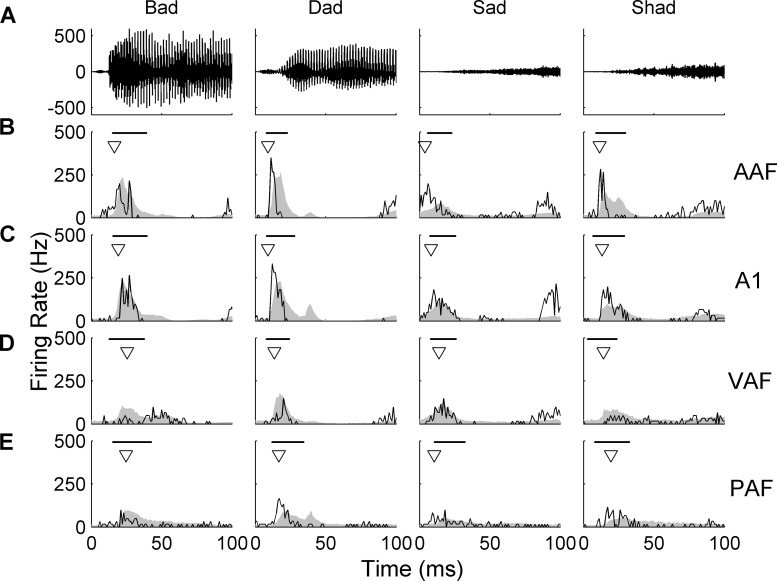
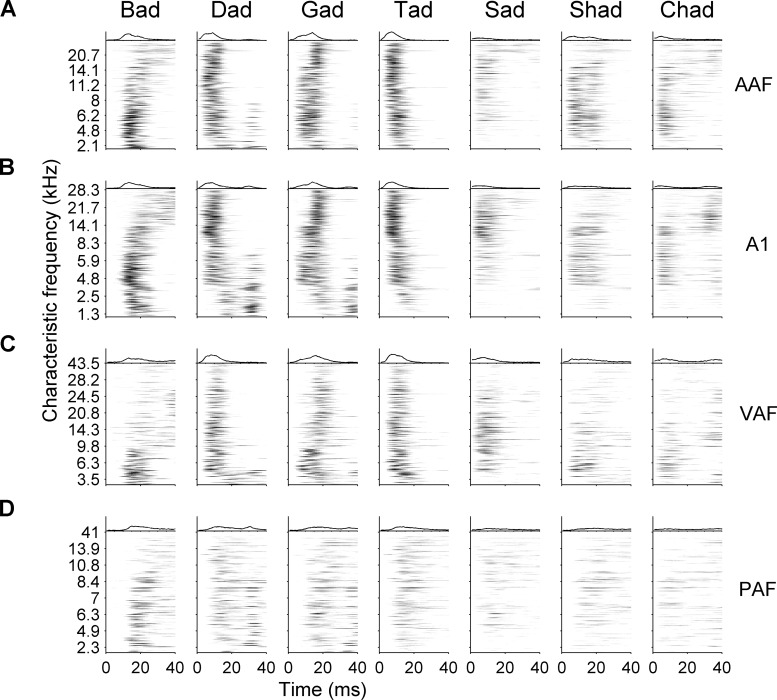
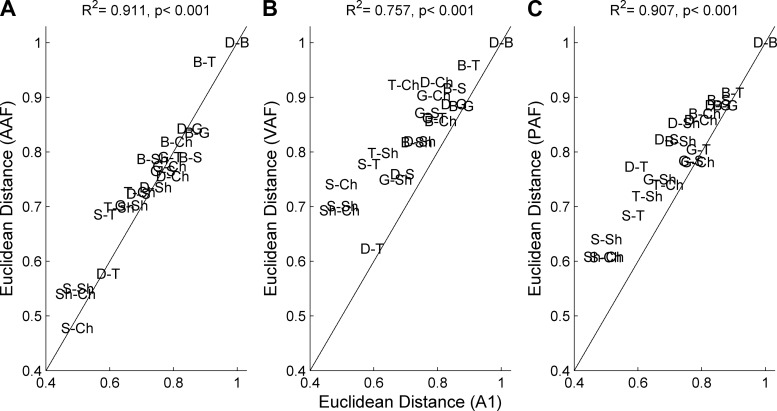
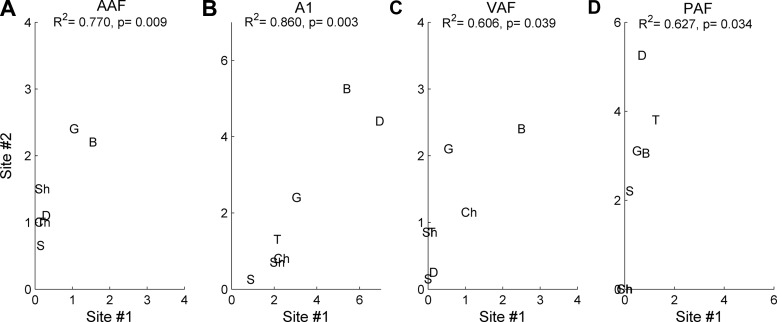
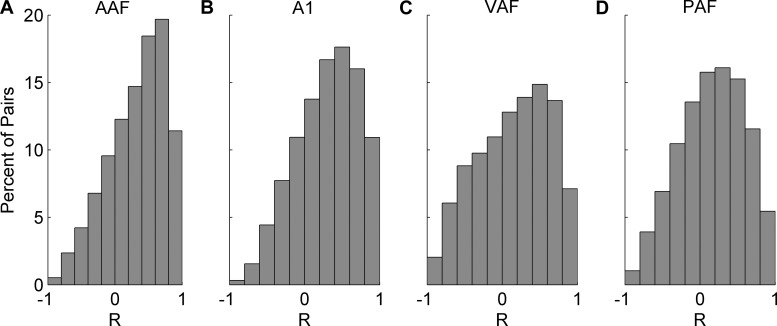
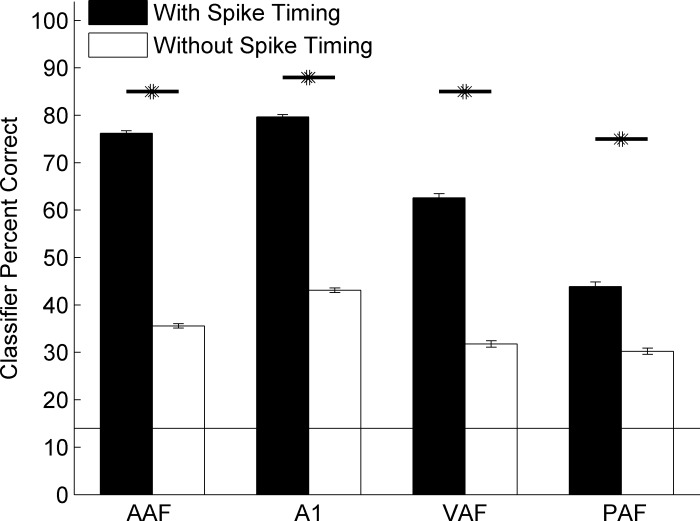
Different speech sounds evoke unique patterns of activity in primary auditory cortex (A1). Behavioral discrimination by rats is well correlated with the distinctness of the A1 patterns evoked by individual consonants, but only when precise spike timing is preserved. In this study we recorded the speech-evoked responses in the primary, anterior, ventral, and posterior auditory fields of the rat and evaluated whether activity in these fields is better correlated with speech discrimination ability when spike timing information is included or eliminated. Spike timing information improved consonant discrimination in all four of the auditory fields examined. Behavioral discrimination was significantly correlated with neural discrimination in all four auditory fields. The diversity of speech responses across recordings sites was greater in posterior and ventral auditory fields compared with A1 and anterior auditor fields. These results suggest that, while the various auditory fields of the rat process speech sounds differently, neural activity in each field could be used to distinguish between consonant sounds with accuracy that closely parallels behavioral discrimination. Earlier observations in the visual and somatosensory systems that cortical neurons do not rely on spike timing should be reevaluated with more complex natural stimuli to determine whether spike timing contributes to sensory encoding.
Keywords: auditory cortex; diversity; parallel hierarchy; rat; spike timing.
Figures
References
-
- Ahissar E, Sosnik R, Haidarliu S. Transformation from temporal to rate coding in a somatosensory thalamocortical pathway. Nature 406: 302–306, 2000 - PubMed
-
- Anderson S, Kilgard M, Sloan A, Rennaker R. Response to broadband repetitive stimuli in auditory cortex of the unanesthetized rat. Hear Res 213: 107–117, 2006 - PubMed
-
- Buonomano DV, Merzenich MM. Temporal information transformed into a spatial code by a neural network with realistic properties. Science 267: 1028–1030, 1995 - PubMed
-
- Carrasco A, Lomber SG. Neuronal activation times to simple, complex, and natural sounds in cat primary and nonprimary auditory cortex. J Neurophysiol 106: 1166–1178, 2011 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources