

Crassulacean acid metabolism-cycling in Euphorbia milii
- PMID: 23596548
- PMCID: PMC3628315
- DOI: 10.1093/aobpla/plt014
Crassulacean acid metabolism-cycling in Euphorbia milii
Abstract
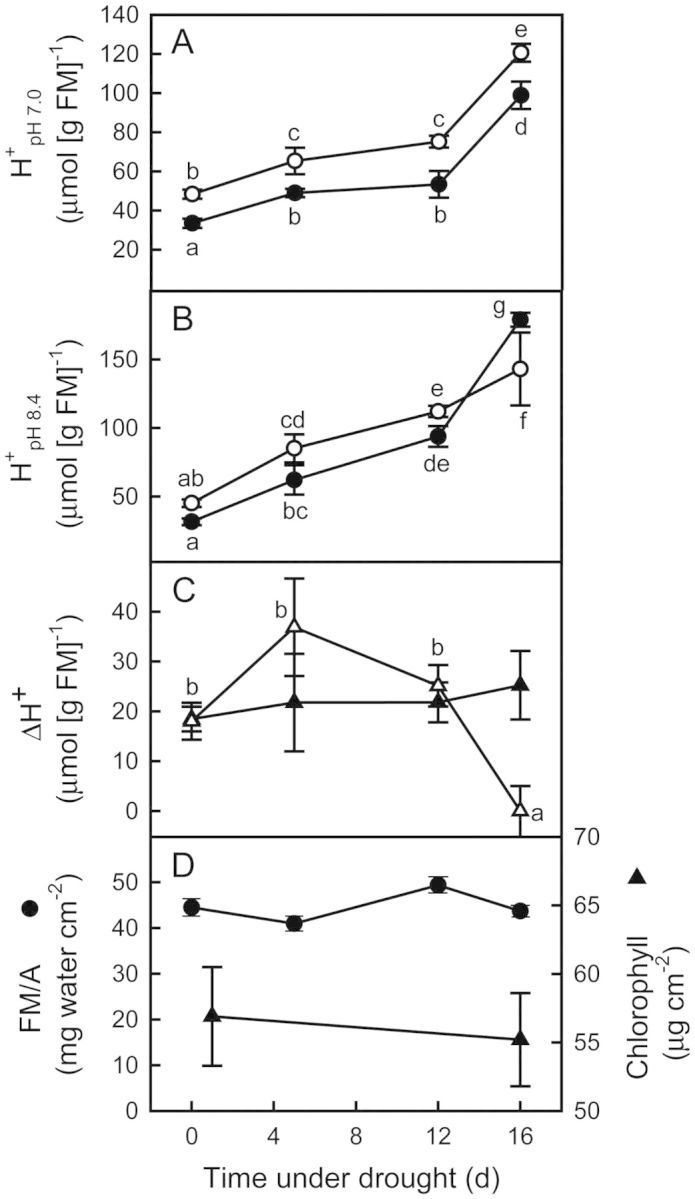
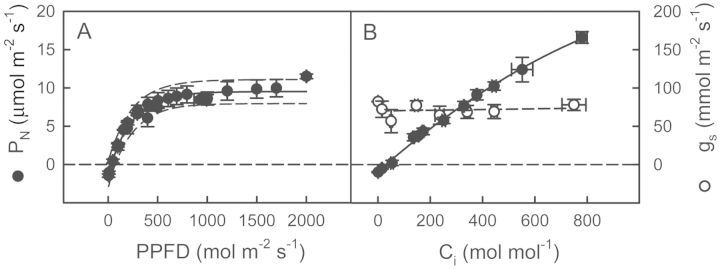
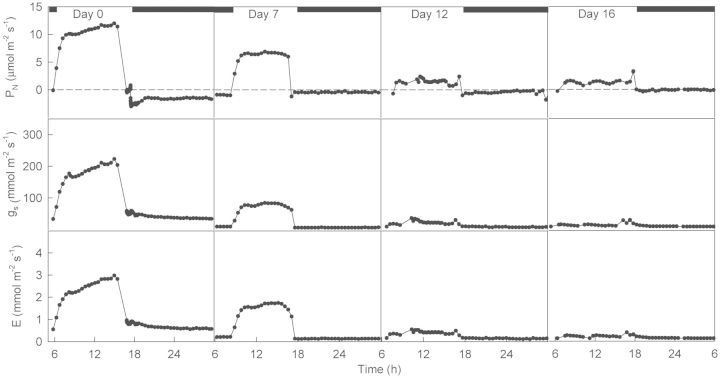
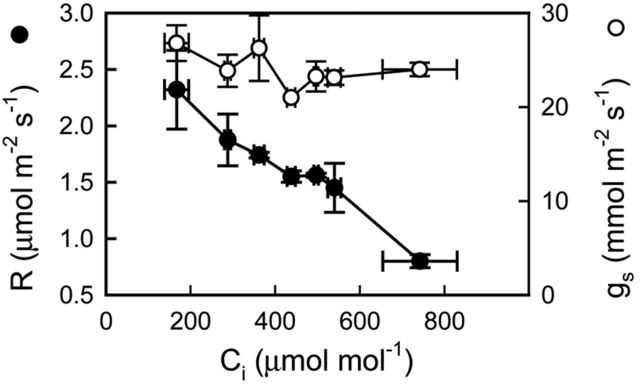
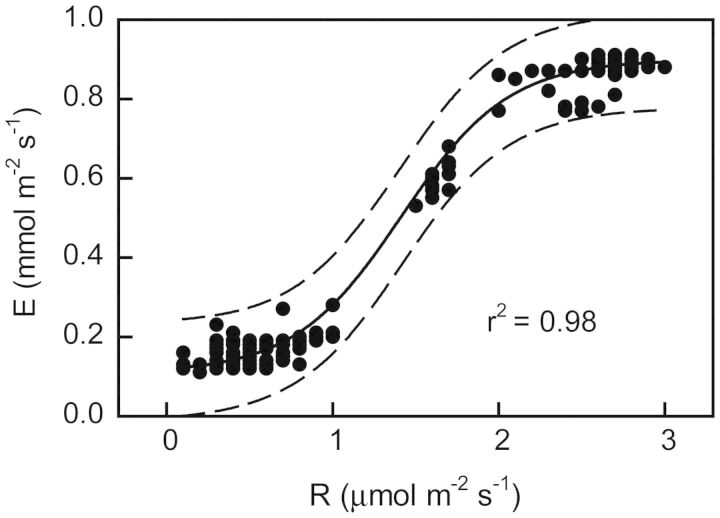
Crassulacean acid metabolism (CAM) occurs in many Euphorbiaceae, particularly Euphorbia, a genus with C3 and C4 species as well. With the aim of contributing to our knowledge of the evolution of CAM in this genus, this study examined the possible occurrence of CAM in Euphorbia milii, a species with leaf succulence and drought tolerance suggestive of this carbon fixation pathway. Leaf anatomy consisted of a palisade parenchyma, a spongy parenchyma and a bundle sheath with chloroplasts, which indicates the possible functioning of C2 photosynthesis. No evidence of nocturnal CO2 fixation was found in plants of E. milii either watered or under drought; watered plants had a low nocturnal respiration rate (R). After 12 days without watering, the photosynthetic rate (P N) decreased 85 % and nocturnal R was nearly zero. Nocturnal H(+) accumulation (ΔH(+)) in watered plants was 18 ± 2 (corresponding to malate) and 18 ± 4 (citrate) μmol H(+) (g fresh mass)(-1). Respiratory CO2 recycling through acid synthesis contributed to a night-time water saving of 2 and 86 % in watered plants and plants under drought, respectively. Carbon isotopic composition (δ(13)C) was -25.2 ± 0.7 ‰ in leaves and -24.7 ± 0.1 ‰ in stems. Evidence was found for the operation of weak CAM in E. milii, with statistically significant ΔH(+), no nocturnal CO2 uptake and values of δ(13)C intermediate between C3 and constitutive CAM plants; ΔH(+) was apparently attributable to both malate and citrate. The results suggest that daily malate accumulation results from recycling of part of the nocturnal respiratory CO2, which helps explain the occurrence of an intermediate value of leaf δ(13)C. Euphorbia milii can be considered as a CAM-cycling species. The significance of the operation of CAM-cycling in E. milii lies in water conservation, rather than carbon acquisition. The possible occurrence of C2 photosynthesis merits research.
Keywords: CAM-cycling; citrate; transpiration; water saving; water-use efficiency.
Figures

Similar articles
-
Leaf anatomical traits which accommodate the facultative engagement of crassulacean acid metabolism in tropical trees of the genus Clusia.J Exp Bot. 2014 Jul;65(13):3513-23. doi: 10.1093/jxb/eru022. Epub 2014 Feb 7. J Exp Bot. 2014. PMID: 24510939
-
Interactive effects of photon fluence rates and drought on CAM-cycling in Delosperma tradescantioides (Mesembryanthemaceae).Physiol Plant. 1998 Jan;102(1):148-154. doi: 10.1034/j.1399-3054.1998.1020119.x. Physiol Plant. 1998. PMID: 35359119
-
CAM photosynthesis in Bulnesia retama (Zygophyllaceae), a non-succulent desert shrub from South America.Ann Bot. 2023 Nov 25;132(4):655-670. doi: 10.1093/aob/mcad114. Ann Bot. 2023. PMID: 37625031 Free PMC article.
-
Achievable productivities of certain CAM plants: basis for high values compared with C3 and C4 plants.New Phytol. 1991 Oct;119(2):183-205. doi: 10.1111/j.1469-8137.1991.tb01022.x. New Phytol. 1991. PMID: 33874131 Review.
-
Crassulacean acid metabolism and fitness under water deficit stress: if not for carbon gain, what is facultative CAM good for?Ann Bot. 2009 Feb;103(4):645-53. doi: 10.1093/aob/mcn145. Epub 2008 Aug 15. Ann Bot. 2009. PMID: 18708641 Free PMC article. Review.
Cited by
-
Uptake and degradation of trimethylamine by Euphorbia milii.Environ Sci Pollut Res Int. 2016 Sep;23(17):17067-76. doi: 10.1007/s11356-016-6874-z. Epub 2016 May 21. Environ Sci Pollut Res Int. 2016. PMID: 27209635
-
Organic Acids: The Pools of Fixed Carbon Involved in Redox Regulation and Energy Balance in Higher Plants.Front Plant Sci. 2016 Jul 15;7:1042. doi: 10.3389/fpls.2016.01042. eCollection 2016. Front Plant Sci. 2016. PMID: 27471516 Free PMC article. Review.
-
Laying the Foundation for Crassulacean Acid Metabolism (CAM) Biodesign: Expression of the C4 Metabolism Cycle Genes of CAM in Arabidopsis.Front Plant Sci. 2019 Feb 11;10:101. doi: 10.3389/fpls.2019.00101. eCollection 2019. Front Plant Sci. 2019. PMID: 30804970 Free PMC article.
-
C4 trees have a broader niche than their close C3 relatives.J Exp Bot. 2022 May 23;73(10):3189-3204. doi: 10.1093/jxb/erac113. J Exp Bot. 2022. PMID: 35293994 Free PMC article.
-
Responses of Leaf Anatomy and CO2 Concentrating Mechanisms of the Aquatic Plant Ottelia cordata to Variable CO2.Front Plant Sci. 2020 Aug 14;11:1261. doi: 10.3389/fpls.2020.01261. eCollection 2020. Front Plant Sci. 2020. PMID: 32922428 Free PMC article.
References
-
- Batanouny KH, Stichler W, Ziegler H. Photosynthetic pathways and ecological distribution of Euphorbia species in Egypt. Oecologia. 1991;87:565–569. - PubMed
-
- Borland AM, Griffiths H, Maxwell C, Broadmeadow MSJ, Griffiths NM, Barnes JD. On the ecophysiology of the Clusiaceae in Trinidad: expression of CAM in Clusia minor L. during the transition from wet to dry season and characterization of tree endemic species. New Phytologist. 1992;122:349–357. - PubMed
-
- Bruinsma JB. The quantitative analysis of chlorophylls a and b in plant extracts. Photochemistry and Photobiology. 1963;2:241–249.
-
- Busch FA, Sage TL, Cousins AB, Sage RF. C3 plants enhance rates of photosynthesis by reassimilating photorespired and respired CO2. Plant, Cell and Environment. 2013;36:200–212. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
