

Suppression of plant defense responses by extracellular metabolites from Pseudomonas syringae pv. tabaci in Nicotiana benthamiana
- PMID: 23597256
- PMCID: PMC3648423
- DOI: 10.1186/1471-2229-13-65
Suppression of plant defense responses by extracellular metabolites from Pseudomonas syringae pv. tabaci in Nicotiana benthamiana
Abstract
Background: Pseudomonas syringae pv. tabaci (Pstab) is the causal agent of wildfire disease in tobacco plants. Several pathovars of Pseudomonas syringae produce a phytotoxic extracellular metabolite called coronatine (COR). COR has been shown to suppress plant defense responses. Interestingly, Pstab does not produce COR but still actively suppresses early plant defense responses. It is not clear if Pstab produces any extracellular metabolites that actively suppress early defense during bacterial pathogenesis.
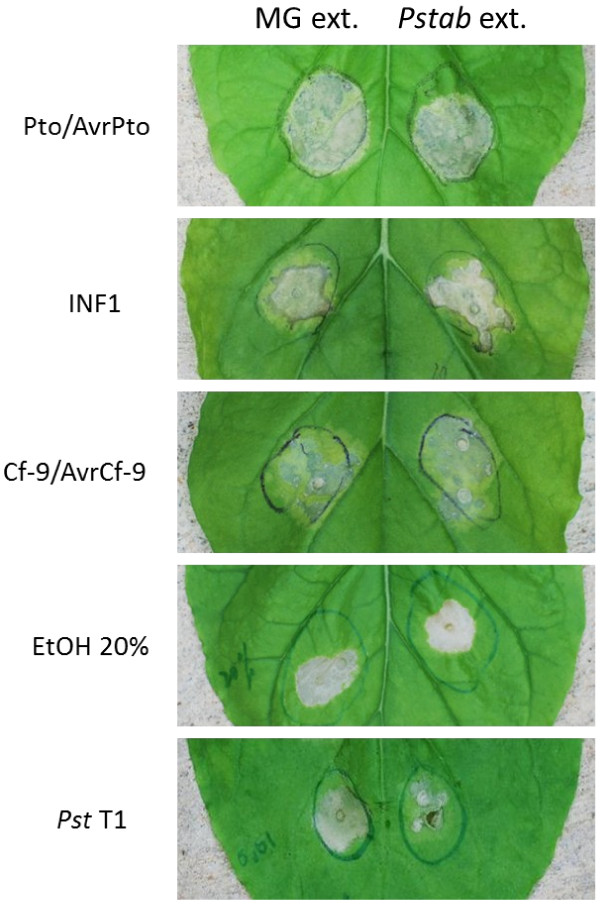
Results: We found that the Pstab extracellular metabolite extracts (Pstab extracts) remarkably suppressed stomatal closure and nonhost hypersensitive response (HR) cell death induced by a nonhost pathogen, P. syringae pv. tomato T1 (Pst T1), in Nicotiana benthamiana. We also found that the accumulation of nonhost pathogens, Pst T1 and P. syringae pv. glycinea (Psgly), was increased in N. benthamiana plants upon treatment with Pstab extracts . The HR cell death induced by Pathogen-Associated Molecular Pattern (INF1), gene-for-gene interaction (Pto/AvrPto and Cf-9/AvrCf-9) and ethanol was not delayed or suppressed by Pstab extracts. We performed metabolite profiling to investigate the extracellular metabolites from Pstab using UPLC-qTOF-MS and identified 49 extracellular metabolites from the Pstab supernatant culture. The results from gene expression profiling of PR-1, PR-2, PR-5, PDF1.2, ABA1, COI1, and HSR203J suggest that Pstab extracellular metabolites may interfere with SA-mediated defense pathways.
Conclusions: In this study, we found that Pstab extracts suppress plant defense responses such as stomatal closure and nonhost HR cell death induced by the nonhost bacterial pathogen Pst T1 in N. benthamiana.
Figures





References
-
- Hirano SS, Upper CD. Population biology and epidemiology of Pseudomonas syringae. Annu Rev Phytopathol. 1990;28:155–177. doi: 10.1146/annurev.py.28.090190.001103. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
