

Not all MSCs can act as pericytes: functional in vitro assays to distinguish pericytes from other mesenchymal stem cells in angiogenesis
- PMID: 23600480
- PMCID: PMC3749721
- DOI: 10.1089/scd.2012.0415
Not all MSCs can act as pericytes: functional in vitro assays to distinguish pericytes from other mesenchymal stem cells in angiogenesis
Abstract
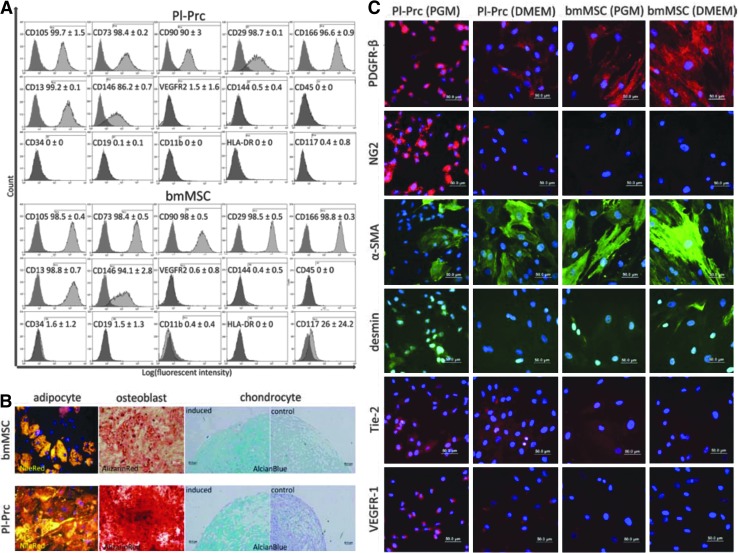
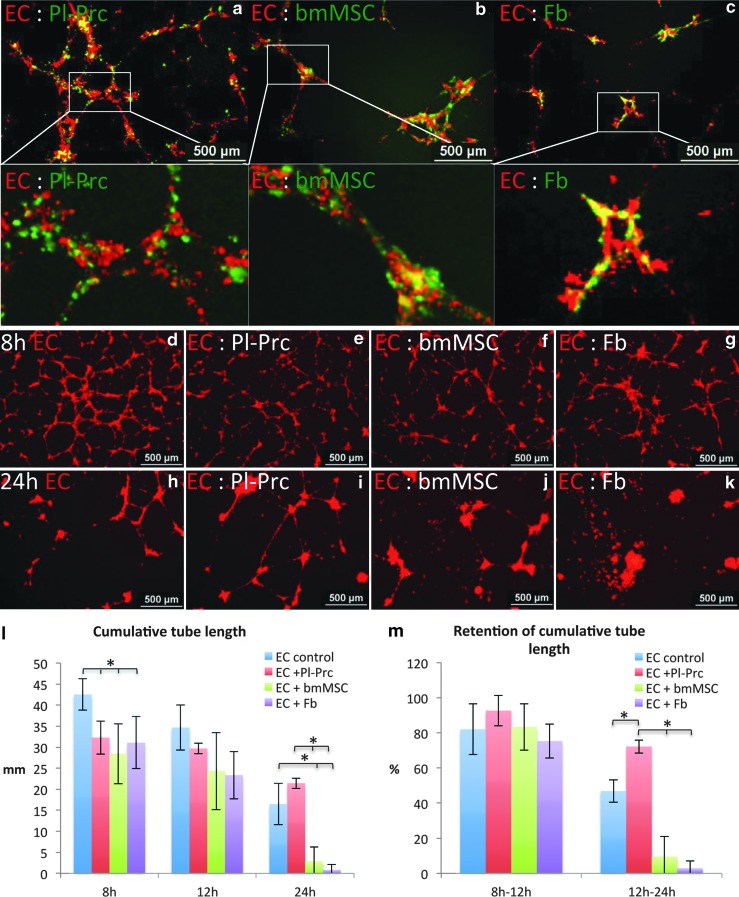
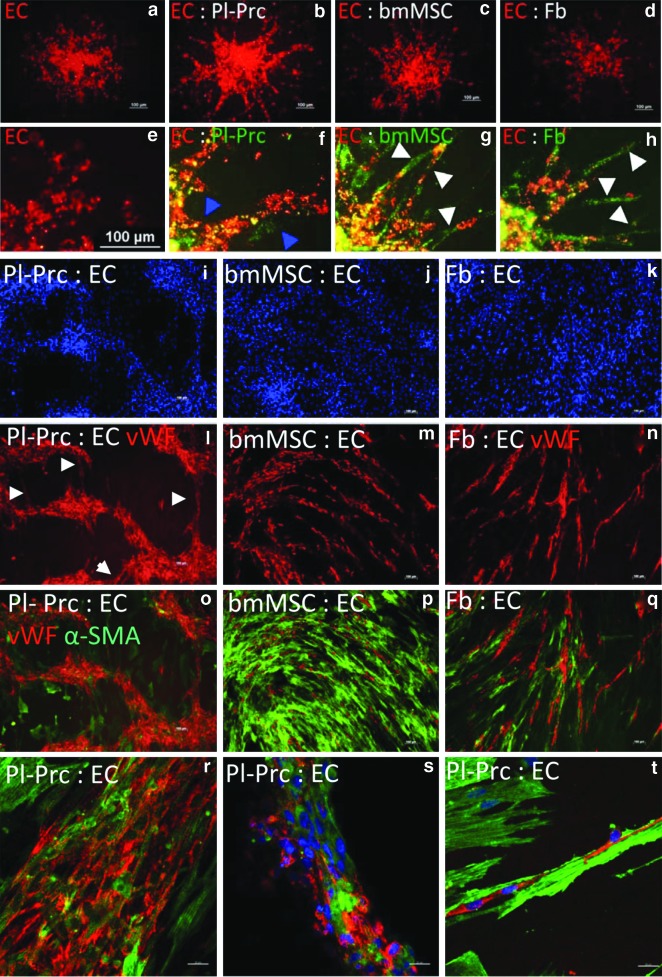
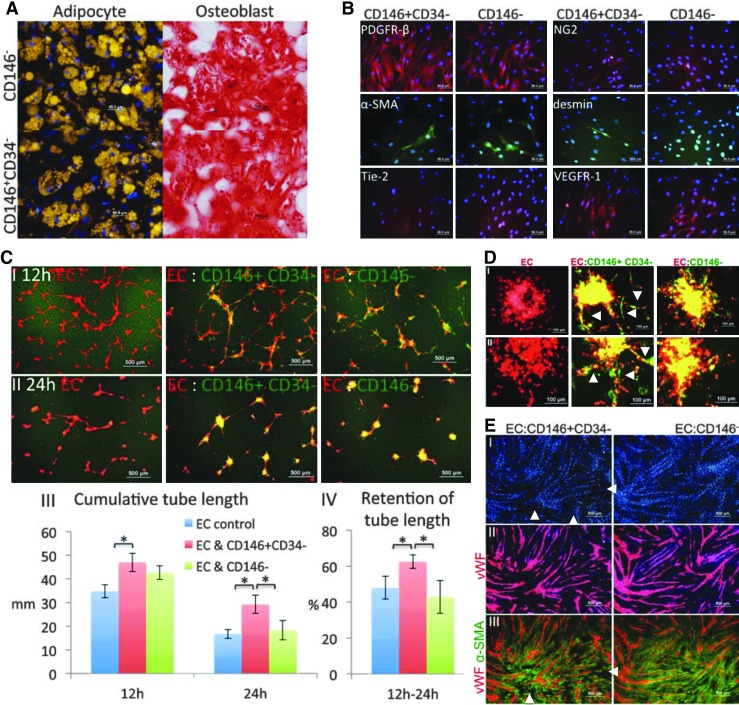
Pericytes play a crucial role in angiogenesis and vascular maintenance. They can be readily identified in vivo and isolated as CD146(+)CD34(-) cells from various tissues. Whether these and other markers reliably identify pericytes in vitro is unclear. CD146(+)CD34(-) selected cells exhibit multilineage potential. Thus, their perivascular location might represent a stem cell niche. This has spurred assumptions that not only all pericytes are mesenchymal stromal cells (MSCs), but also that all MSCs can act as pericytes. Considering this hypothesis, we developed functional assays by confronting test cells with endothelial cultures based on matrigel assay, spheroid sprouting, and cord formation. We calibrated these assays first with commercial cell lines [CD146(+)CD34(-) placenta-derived pericytes (Pl-Prc), bone marrow (bm)MSCs and fibroblasts]. We then functionally compared the angiogenic abilities of CD146(+)CD34(-)selected bmMSCs with CD146(-) selected bmMSCs from fresh human bm aspirates. We show here that only CD146(+)CD34(-) selected Pl-Prc and CD146(+)CD34(-) selected bmMSCs maintain endothelial tubular networks on matrigel and improve endothelial sprout morphology. CD146(-) selected bmMSCs neither showed these abilities, nor did they attain pericyte function despite progressive CD146 expression once passaged. Thus, cell culture conditions appear to influence expression of this and other reported pericyte markers significantly without correlation to function. The newly developed assays, therefore, promise to close a gap in the in vitro identification of pericytes via function. Indeed, our functional data suggest that pericytes represent a subpopulation of MSCs in bm with a specialized role in vascular biology. However, these functions are not inherent to all MSCs.
Figures
References
-
- Sims DE. The pericyte—a review. Tissue Cell. 1986;18:153–174. - PubMed
-
- Armulik A. Genové G. Betsholtz C. Pericytes: developmental, physiological, and pathological perspectives, problems, and promises. Dev Cell. 2011;21:193–215. - PubMed
-
- Darland DC. D'Amore PA. TGF beta is required for the formation of capillary-like structures in three-dimensional cocultures of 10T1/2 and endothelial cells. Angiogenesis. 2001;4:11–20. - PubMed
-
- Dar A. Domev H. Ben-Yosef O. Tzukerman M. Zeevi-Levin N. Novak A, et al. Multipotent vasculogenic pericytes from human pluripotent stem cells promote recovery of murine ischemic limb. Circulation. 2012;125:87–99. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
