

doi: 10.1016/j.pt.2013.03.006.
Epub 2013 Apr 16.
The iron link between malaria and invasive non-typhoid Salmonella infections
Affiliations
- PMID: 23601932
- PMCID: PMC4521076
- DOI: 10.1016/j.pt.2013.03.006
Item in Clipboard
The iron link between malaria and invasive non-typhoid Salmonella infections
Trends Parasitol.
2013 May.
Abstract
Epidemiological studies have demonstrated an association between malaria and invasive non-typhoid Salmonella (NTS) infections, especially in children. We explore the role of iron as a possible cofactor in this association. Malarial disease, among others, is associated with enhanced erythrophagocytosis and inflammation, which increases the iron content of macrophages and thereby also the survival of Salmonella spp. within macrophages. Whether iron supplementation programs augment the risk of invasive NTS infections in malaria-endemic regions is an important global health issue that still needs to be determined.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures
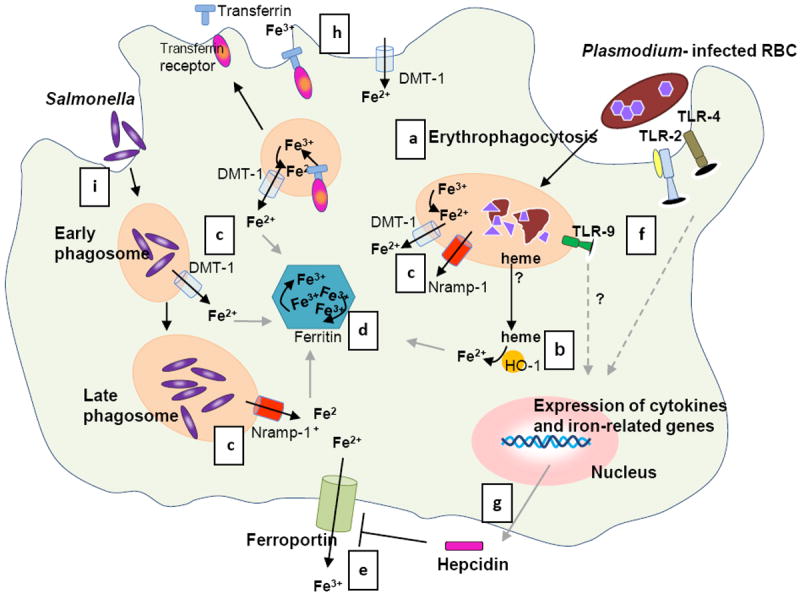
Phagocytosis of both uninfected and infected RBCs (a) is increased during the blood stage malaria infection [83], which results in degradation of the RBCs by proteolytic enzymes into heme. (b) HO-1 converts heme into iron (and carbon monoxide and biliverdin). (c) Excess iron is transported to the cytosol via phagosomal transporters DMT-1 and Nramp-1 [24] and further processed: (i) stored in ferritin (d), and (ii) used in metabolic processes or released from the cell via ferroportin (e) [26, 79]. (f) Meanwhile, parasite products activate the innate immune system via Toll Like receptors (TLR) 2, 4 and 9 [84]. This systemic response during malaria induces hepatic hepcidin production; (e) hepcidin functions by blocking ferroportin [29, 78]. In addition, monocytes and macrophages also express hepcidin upon stimulation with various pro-inflammatory cytokines and parasitized RBCs [36-38], (g) which may result in autocrine ferroportin blocking. (h) In addition, inflammatory stimuli inhibit ferroportin and modulate cellular iron uptake by DMT-1 and transferrin receptor [36, 39]. As a consequence of these processes iron is sequestered in macrophages. (i)
Salmonella enters the cell via endocytosis and proliferates in phagosomes. Nramp-1 expression is required to control Salmonella growth by depleting the phagosome of iron (c) [46]. In a co-infection, Salmonellae spp. may benefit from the increased cellular iron induced by a malaria infection and establish an infection. Whether both pathogens reside in the same macrophage during invasive NTS infection and malaria as depicted in the figure is unknown. Illustration by A. Kartikasari.
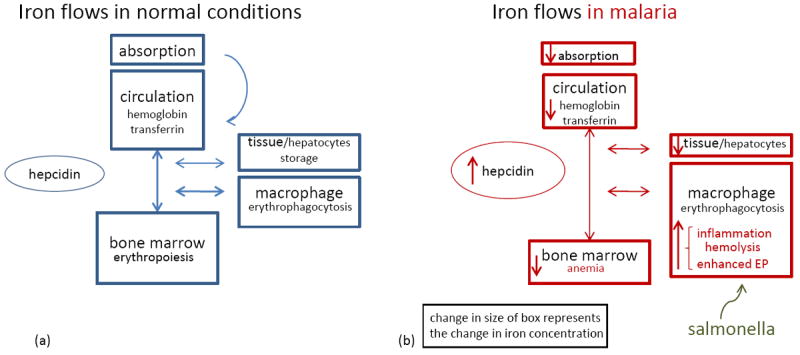
(a) A schematic presentation of iron flows in normal circumstances. Hepcidin controls the amount of iron absorbed from the diet and the release of iron from macrophages from the reticulo-endothelial system [29]. There is a steady state of iron recycling from senescent RBCs that are degraded in macrophages into iron. This iron is transported via transferrin in the circulation towards the bone marrow where iron is essential for erythropoiesis. Body iron losses are minimal and not regulated. (b) Iron flows in malaria. During malaria infection the body iron homeostasis changes, but total amount of body iron remains similar as is visualized by the change in the size of boxes. Inflammatory factors increase hepcidin release [30, 33, 76, 77]. As a consequence, absorption of iron from the diet is impaired, iron is redistributed to macrophages, less iron is bound to transferrin and iron stores become depleted. Finally, the erythropoiesis is impaired, due to hepcidin-mediated iron restriction, in addition to malaria-specific inhibitory factors (e.g., cytokines, hemozoin) [71]. Also, a blood stage malaria infection is hallmarked by hemolysis and increased phagocytosis of parasitized and non infected RBCs, which also augments the macrophage iron content. As suggested, the increased iron availability in malaria could facilitate the growth and replication of Salmonellae spp. Abbreviation: EP, erythrophagocytosis.
Similar articles
-
Malaria parasite infection compromises control of concurrent systemic non-typhoidal Salmonella infection via IL-10-mediated alteration of myeloid cell function.PLoS Pathog. 2014 May 1;10(5):e1004049. doi: 10.1371/journal.ppat.1004049. eCollection 2014 May. PLoS Pathog. 2014. PMID: 24787713 Free PMC article.
-
Immunological bases of increased susceptibility to invasive nontyphoidal Salmonella infection in children with malaria and anaemia.Microbes Infect. 2018 Oct-Nov;20(9-10):589-598. doi: 10.1016/j.micinf.2017.11.014. Epub 2017 Dec 15. Microbes Infect. 2018. PMID: 29248635 Free PMC article. Review.
-
Loss of Humoral and Cellular Immunity to Invasive Nontyphoidal Salmonella during Current or Convalescent Plasmodium falciparum Infection in Malawian Children.Clin Vaccine Immunol. 2017 Jul 5;24(7):e00057-17. doi: 10.1128/CVI.00057-17. Print 2017 Jul. Clin Vaccine Immunol. 2017. PMID: 28515136 Free PMC article.
-
Infectious disease: Opposing effects of IL-10.Nat Rev Immunol. 2014 Jun;14(6):356. doi: 10.1038/nri3693. Nat Rev Immunol. 2014. PMID: 24854586 No abstract available.
-
The association between malaria and non-typhoid Salmonella bacteraemia in children in sub-Saharan Africa: a literature review.Malar J. 2014 Oct 13;13:400. doi: 10.1186/1475-2875-13-400. Malar J. 2014. PMID: 25311375 Free PMC article. Review.
Cited by
-
Epidemic increase in Salmonella bloodstream infection in children, Bwamanda, the Democratic Republic of Congo.Eur J Clin Microbiol Infect Dis. 2014 Jan;33(1):79-87. doi: 10.1007/s10096-013-1931-8. Epub 2013 Aug 24. Eur J Clin Microbiol Infect Dis. 2014. PMID: 23975545
-
Role of Activins in Hepcidin Regulation during Malaria.Infect Immun. 2017 Nov 17;85(12):e00191-17. doi: 10.1128/IAI.00191-17. Print 2017 Dec. Infect Immun. 2017. PMID: 28893916 Free PMC article.
-
Characterization of the Invasive, Multidrug Resistant Non-typhoidal Salmonella Strain D23580 in a Murine Model of Infection.PLoS Negl Trop Dis. 2015 Jun 19;9(6):e0003839. doi: 10.1371/journal.pntd.0003839. eCollection 2015 Jun. PLoS Negl Trop Dis. 2015. PMID: 26091096 Free PMC article.
-
Dissemination of non-typhoidal Salmonella during Plasmodium chabaudi infection affects anti-malarial immunity.Parasitol Res. 2019 Jul;118(7):2277-2285. doi: 10.1007/s00436-019-06349-z. Epub 2019 May 23. Parasitol Res. 2019. PMID: 31119381 Free PMC article.
-
Disrupted Iron Metabolism and Mortality during Co-infection with Malaria and an Intestinal Gram-Negative Extracellular Pathogen.Cell Rep. 2021 Jan 12;34(2):108613. doi: 10.1016/j.celrep.2020.108613. Cell Rep. 2021. PMID: 33440153 Free PMC article.
References
-
- McLean E, et al. Worldwide prevalence of anaemia, WHO Vitamin and Mineral Nutrition Information System, 1993-2005. Public Health Nutr. 2009;12:444–454. - PubMed
-
- Sazawal S, et al. Effects of routine prophylactic supplementation with iron and folic acid on admission to hospital and mortality in preschool children in a high malaria transmission setting: community-based, randomised, placebo-controlled trial. Lancet. 2006;367:133–143. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
