

Compartmentalized oxidative stress in dopaminergic cell death induced by pesticides and complex I inhibitors: distinct roles of superoxide anion and superoxide dismutases
- PMID: 23602909
- PMCID: PMC3883883
- DOI: 10.1016/j.freeradbiomed.2013.04.021
Compartmentalized oxidative stress in dopaminergic cell death induced by pesticides and complex I inhibitors: distinct roles of superoxide anion and superoxide dismutases
Abstract
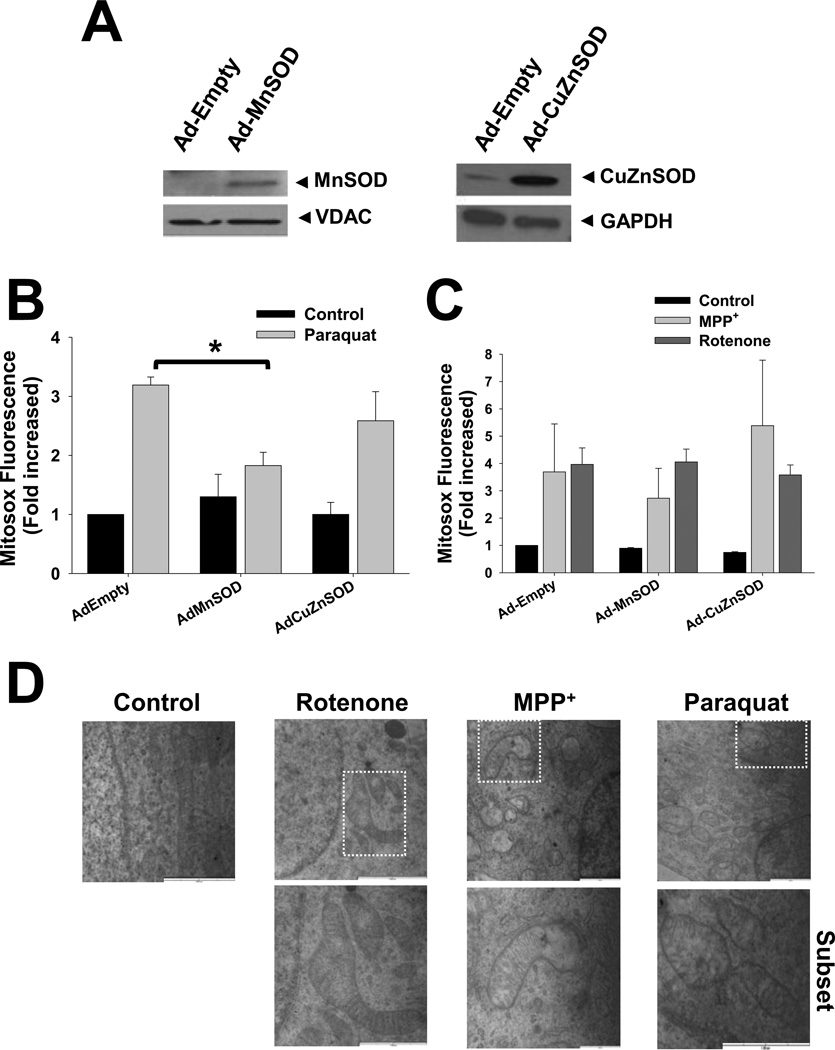
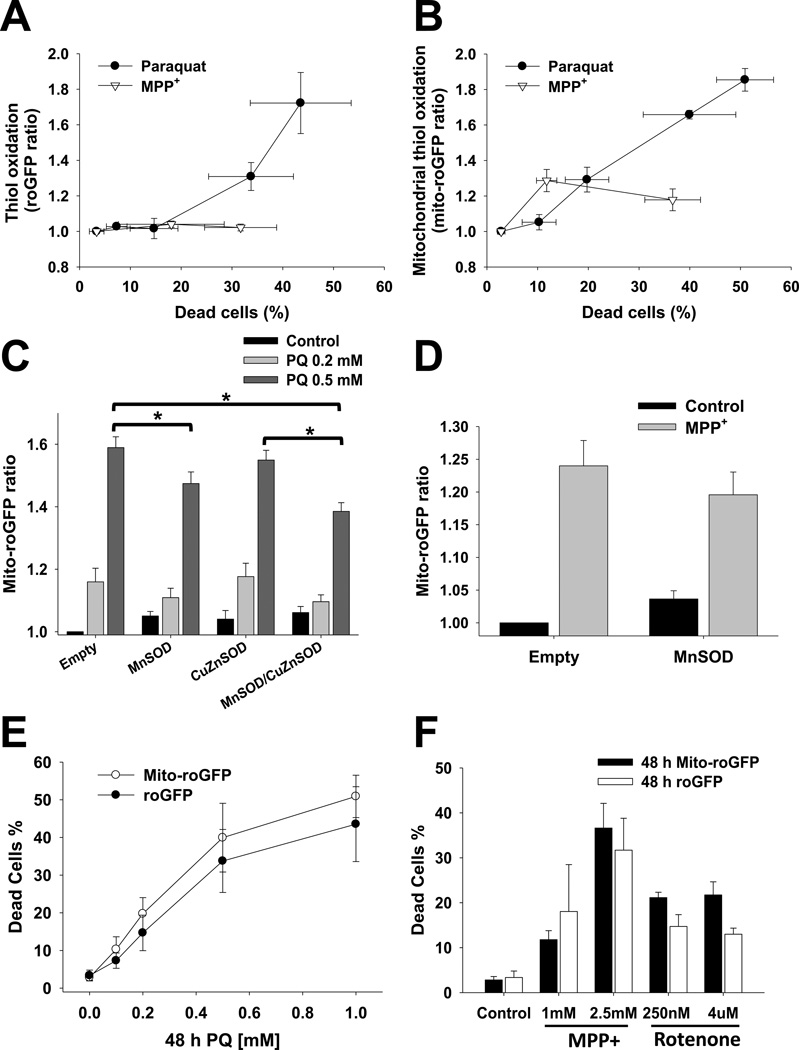
The loss of dopaminergic neurons induced by the parkinsonian toxins paraquat, rotenone, and 1-methyl-4-phenylpyridinium (MPP(+)) is associated with oxidative stress. However, controversial reports exist regarding the source/compartmentalization of reactive oxygen species (ROS) generation and its exact role in cell death. We aimed to determine in detail the role of superoxide anion (O2(•-)), oxidative stress, and their subcellular compartmentalization in dopaminergic cell death induced by parkinsonian toxins. Oxidative stress and ROS formation were determined in the cytosol, intermembrane (IMS), and mitochondrial matrix compartments, using dihydroethidine derivatives and the redox sensor roGFP, as well as electron paramagnetic resonance spectroscopy. Paraquat induced an increase in ROS and oxidative stress in both the cytosol and the mitochondrial matrix prior to cell death. MPP(+) and rotenone primarily induced an increase in ROS and oxidative stress in the mitochondrial matrix. No oxidative stress was detected at the level of the IMS. In contrast to previous studies, overexpression of manganese superoxide dismutase (MnSOD) or copper/zinc SOD (CuZnSOD) had no effect on alterations in ROS steady-state levels, lipid peroxidation, loss of mitochondrial membrane potential (ΔΨm), and dopaminergic cell death induced by MPP(+) or rotenone. In contrast, paraquat-induced oxidative stress and cell death were selectively reduced by MnSOD overexpression, but not by CuZnSOD or manganese-porphyrins. However, MnSOD also failed to prevent ΔΨm loss. Finally, paraquat, but not MPP(+) or rotenone, induced the transcriptional activation of the redox-sensitive antioxidant response elements (ARE) and nuclear factor kappa-B (NF-κB). These results demonstrate a selective role of mitochondrial O2(•-) in dopaminergic cell death induced by paraquat, and show that toxicity induced by the complex I inhibitors rotenone and MPP(+) does not depend directly on mitochondrial O2(•-) formation.
Keywords: CuZnSOD; Environmental; MPP+; MnSOD; Paraquat; Parkinson's disease; Pesticides; Porphyrins; Rotenone; SOD; roGFP.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures





References
-
- Lees AJ. Unresolved issues relating to the shaking palsy on the celebration of James Parkinson's 250th birthday. Mov Disord. 2007;22(Suppl 17):S327–S334. - PubMed
-
- Alam ZI, Daniel SE, Lees AJ, Marsden DC, Jenner P, Halliwell B. A generalised increase in protein carbonyls in the brain in Parkinson's but not incidental Lewy body disease. J Neurochem. 1997;69:1326–1329. - PubMed
-
- Dexter DT, Carter CJ, Wells FR, Javoy-Agid F, Agid Y, Lees A, Jenner P, Marsden CD. Basal lipid peroxidation in substantia nigra is increased in Parkinson's disease. J Neurochem. 1989;52:381–389. - PubMed
-
- Alam ZI, Jenner A, Daniel SE, Lees AJ, Cairns N, Marsden CD, Jenner P, Halliwell B. Oxidative DNA damage in the parkinsonian brain: an apparent selective increase in 8-hydroxyguanine levels in substantia nigra. J Neurochem. 1997;69:1196–1203. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
