

Going forward with genetics: recent technological advances and forward genetics in mice
- PMID: 23608223
- PMCID: PMC3644711
- DOI: 10.1016/j.ajpath.2013.02.002
Going forward with genetics: recent technological advances and forward genetics in mice
Abstract
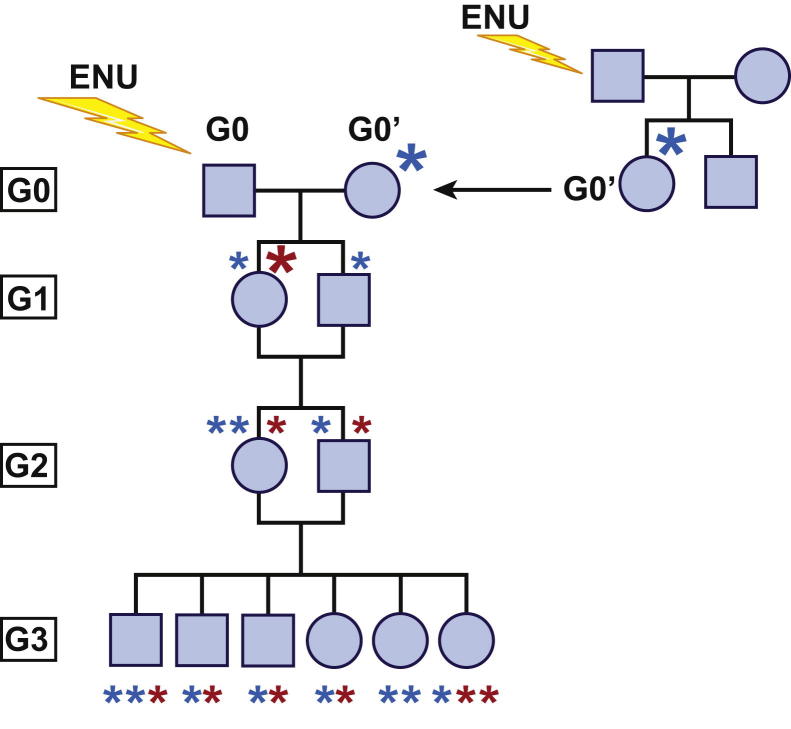
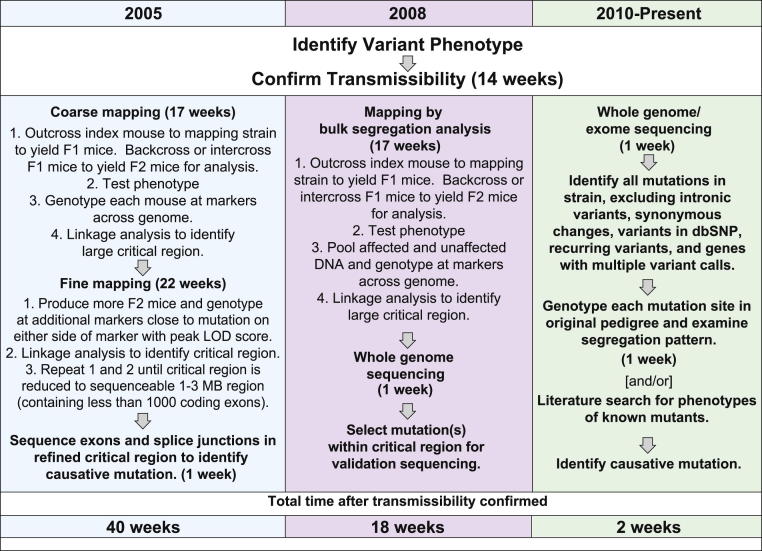
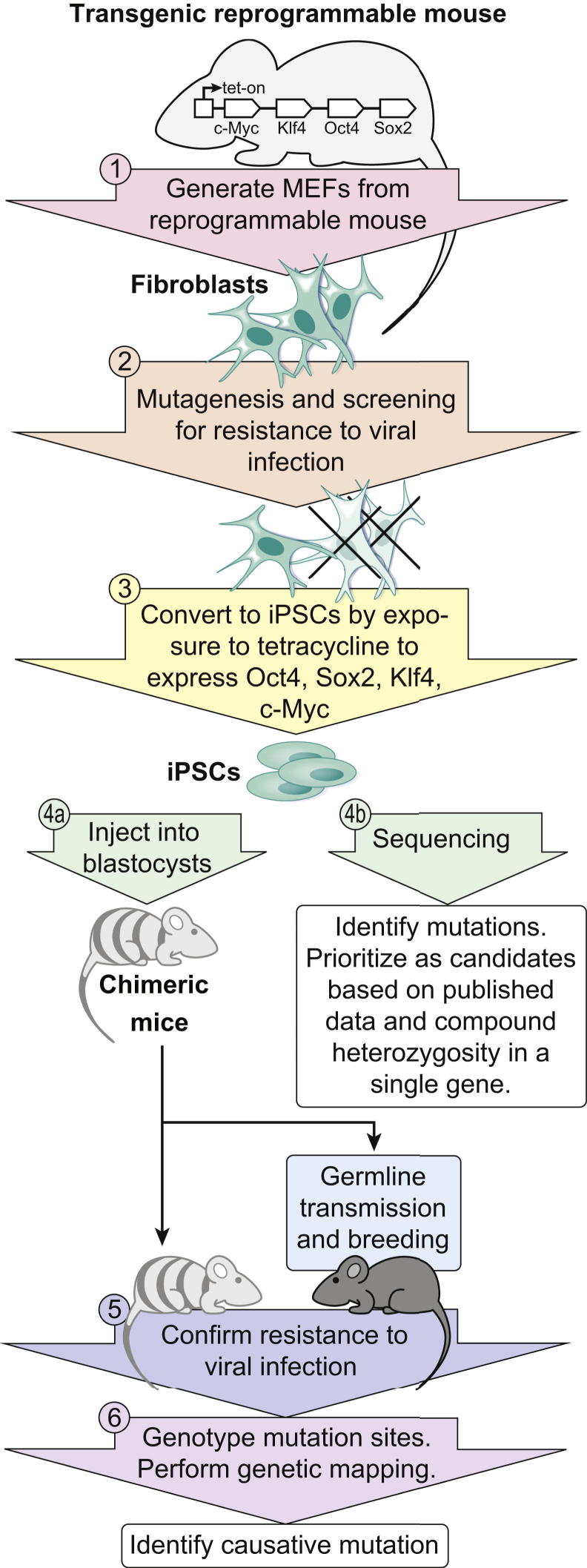
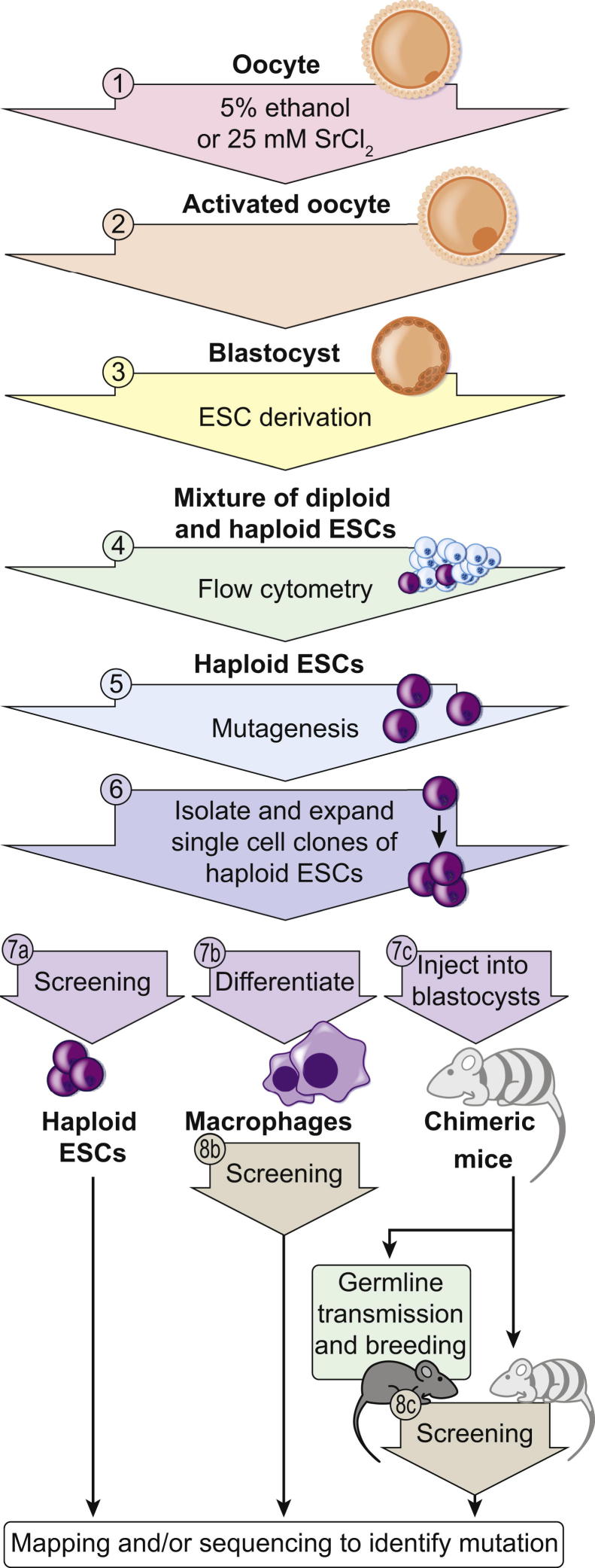
Forward genetic analysis is an unbiased approach for identifying genes essential to defined biological phenomena. When applied to mice, it is one of the most powerful methods to facilitate understanding of the genetic basis of human biology and disease. The speed at which disease-causing mutations can be identified in mutagenized mice has been markedly increased by recent advances in DNA sequencing technology. Creating and analyzing mutant phenotypes may therefore become rate-limiting in forward genetic experimentation. We review the forward genetic approach and its future in the context of recent technological advances, in particular massively parallel DNA sequencing, induced pluripotent stem cells, and haploid embryonic stem cells.
Copyright © 2013 American Society for Investigative Pathology. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Waterston R.H., Lindblad-Toh K., Birney E., Rogers J., Abril J.F., Agarwal P., Mouse Genome Sequencing Consortium Initial sequencing and comparative analysis of the mouse genome. Nature. 2002;420:520–562. - PubMed
-
- Church D.M., Goodstadt L., Hillier L.W., Zody M.C., Goldstein S., She X., Bult C.J., Agarwala R., Cherry J.L., DiCuccio M., Hlavina W., Kapustin Y., Meric P., Maglott D., Birtle Z., Marques A.C., Graves T., Zhou S., Teague B., Potamousis K., Churas C., Place M., Herschleb J., Runnheim R., Forrest D., Amos-Landgraf J., Schwartz D.C., Cheng Z., Lindblad-Toh K., Eichler E.E., Ponting C.P., Mouse Genome Sequencing Consortium Lineage-specific biology revealed by a finished genome assembly of the mouse. PLoS Biol. 2009;7:e1000112. - PMC - PubMed
-
- Skarnes W.C., von Melchner H., Wurst W., Hicks G., Nord A.S., Cox T., Young S.G., Ruiz P., Soriano P., Tessier-Lavigne M., Conklin B.R., Stanford W.L., Rossant J., International Gene Trap Consortium A public gene trap resource for mouse functional genomics. Nat Genet. 2004;36:543–544. - PMC - PubMed
-
- Andrews T.D., Whittle B., Field M.A., Balakishnan B., Zhang Y., Shao Y., Cho V., Kirk M., Singh M., Xia Y., Hager J., Winslade S., Sjollema G., Beutler B., Enders A., Goodnow C.C. Massively parallel sequencing of the mouse exome to accurately identify rare, induced mutations: an immediate source for thousands of new mouse models. Open Biol. 2012;2:120061. - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
