

Sanpodo controls sensory organ precursor fate by directing Notch trafficking and binding γ-secretase
- PMID: 23609534
- PMCID: PMC3639393
- DOI: 10.1083/jcb.201209023
Sanpodo controls sensory organ precursor fate by directing Notch trafficking and binding γ-secretase
Abstract
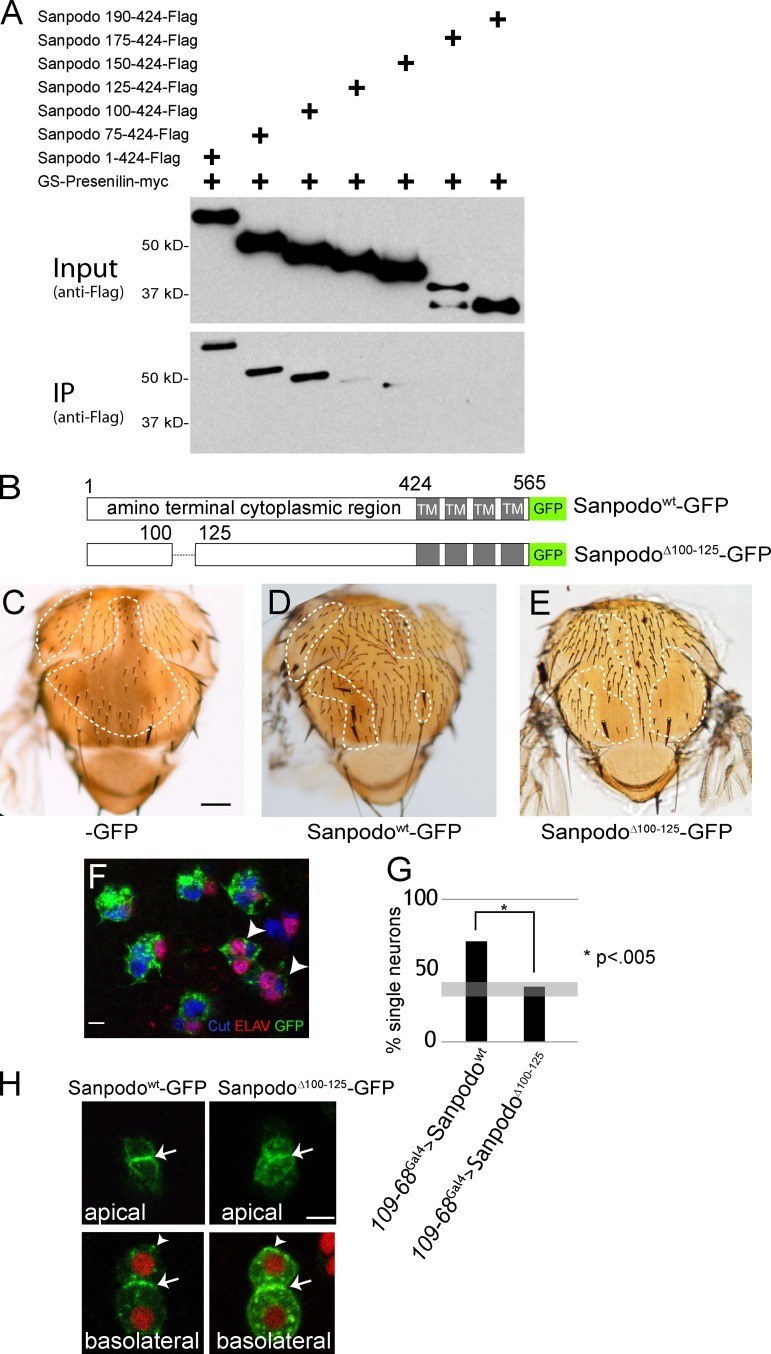
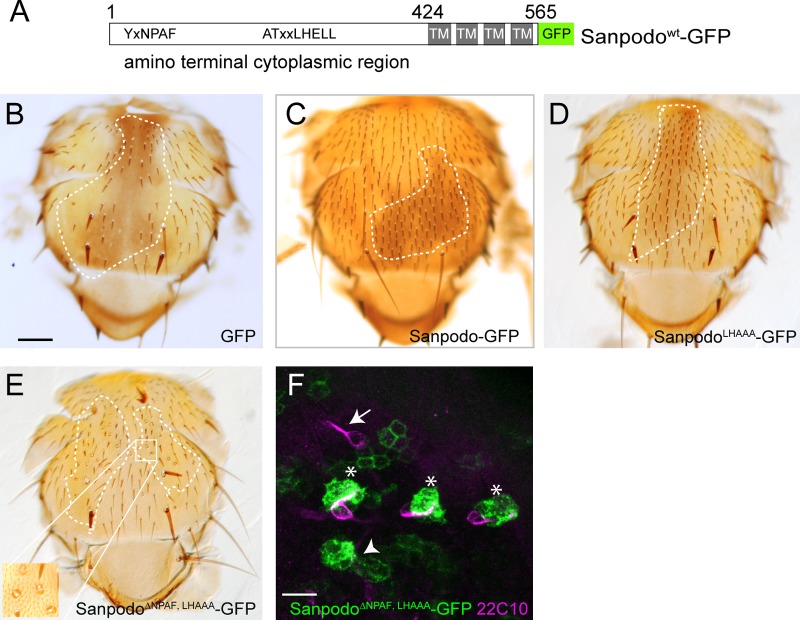
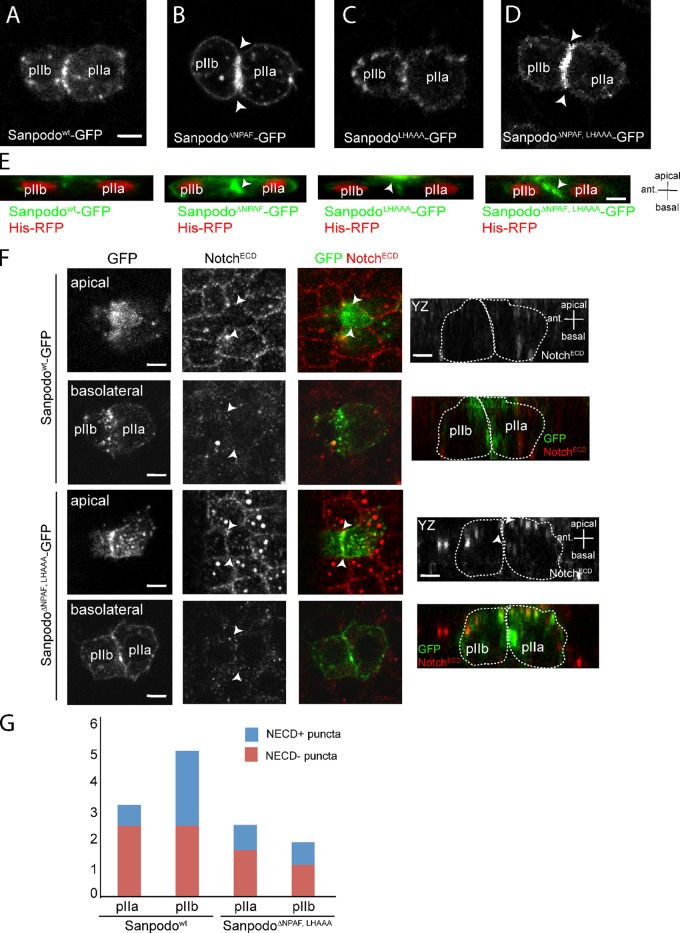
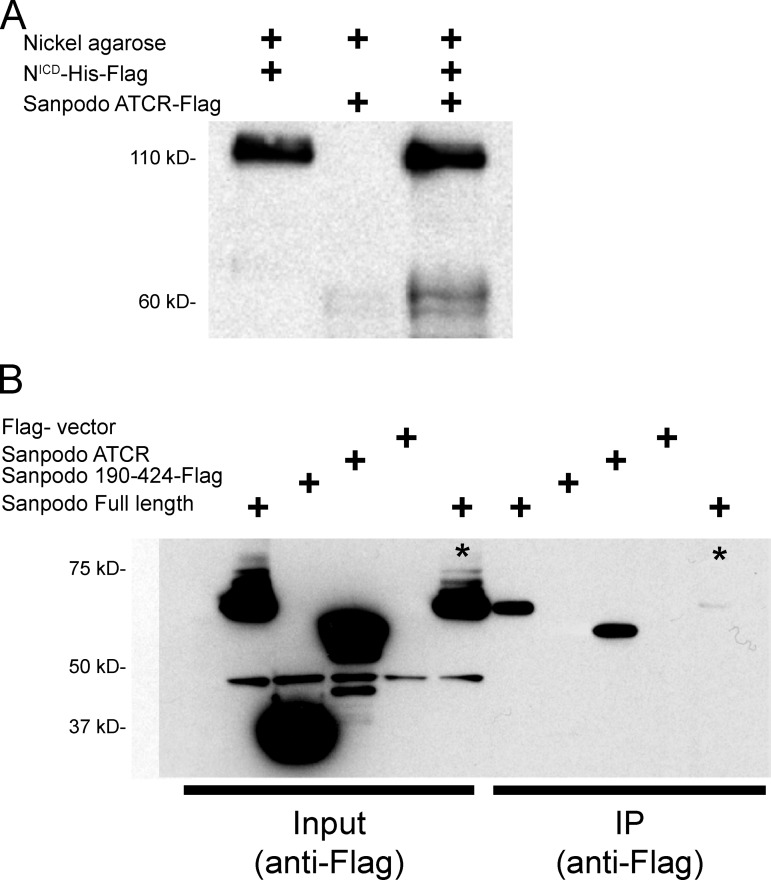
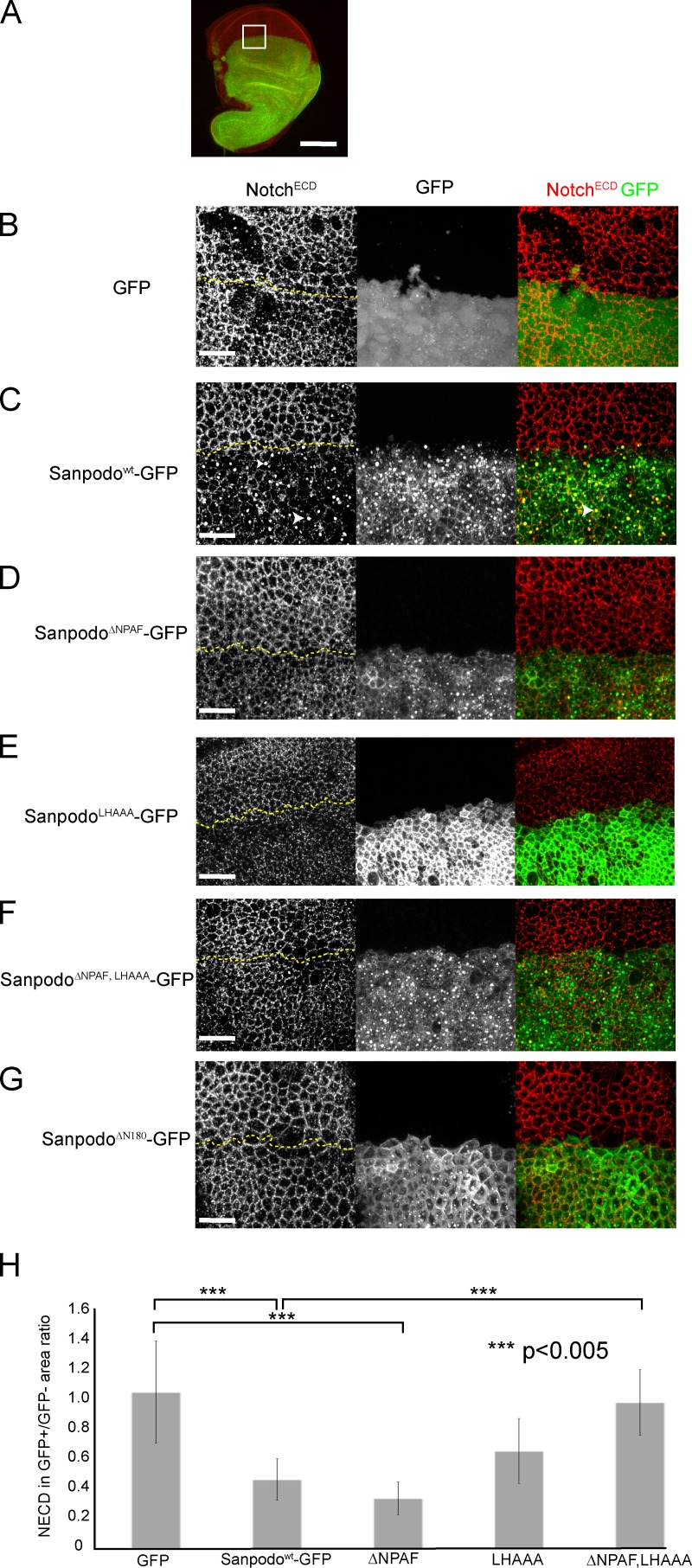
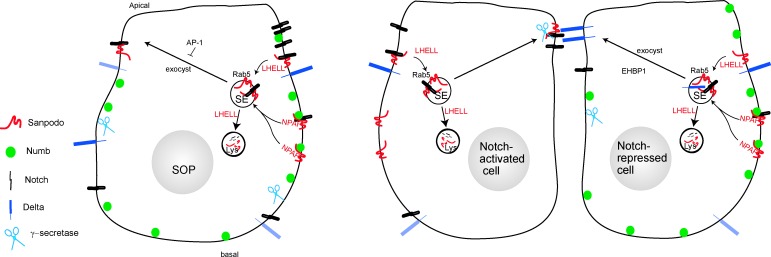
In Drosophila peripheral neurogenesis, Notch controls cell fates in sensory organ precursor (SOP) cells. SOPs undergo asymmetric cell division by segregating Numb, which inhibits Notch signaling, into the pIIb daughter cell after cytokinesis. In contrast, in the pIIa daughter cell, Notch is activated and requires Sanpodo, but its mechanism of action has not been elucidated. As Sanpodo is present in both pIIa and pIIb cells, a second role for Sanpodo in regulating Notch signaling in the low-Notch pIIb cell has been proposed. Here we demonstrate that Sanpodo regulates Notch signaling levels in both pIIa and pIIb cells via distinct mechanisms. The interaction of Sanpodo with Presenilin, a component of the γ-secretase complex, was required for Notch activation and pIIa cell fate. In contrast, Sanpodo suppresses Notch signaling in the pIIb cell by driving Notch receptor internalization. Together, these results demonstrate that a single protein can regulate Notch signaling through distinct mechanisms to either promote or suppress signaling depending on the local cellular context.
Figures
Similar articles
-
Numb and alpha-Adaptin regulate Sanpodo endocytosis to specify cell fate in Drosophila external sensory organs.EMBO Rep. 2005 Sep;6(9):836-42. doi: 10.1038/sj.embor.7400500. EMBO Rep. 2005. PMID: 16113648 Free PMC article.
-
Directional Delta and Notch trafficking in Sara endosomes during asymmetric cell division.Nature. 2009 Apr 23;458(7241):1051-5. doi: 10.1038/nature07854. Epub 2009 Mar 18. Nature. 2009. PMID: 19295516
-
Numb inhibits the recycling of Sanpodo in Drosophila sensory organ precursor.Curr Biol. 2013 Apr 8;23(7):581-7. doi: 10.1016/j.cub.2013.02.020. Epub 2013 Mar 21. Curr Biol. 2013. PMID: 23523246
-
The catalytic core of gamma-secretase: presenilin revisited.Curr Alzheimer Res. 2008 Apr;5(2):147-57. doi: 10.2174/156720508783954677. Curr Alzheimer Res. 2008. PMID: 18393799 Review.
-
Endocytosis and control of Notch signaling.Curr Opin Cell Biol. 2012 Aug;24(4):534-40. doi: 10.1016/j.ceb.2012.06.006. Epub 2012 Jul 18. Curr Opin Cell Biol. 2012. PMID: 22818956 Free PMC article. Review.
Cited by
-
Numb regulates the balance between Notch recycling and late-endosome targeting in Drosophila neural progenitor cells.Mol Biol Cell. 2016 Sep 15;27(18):2857-66. doi: 10.1091/mbc.E15-11-0751. Epub 2016 Jul 27. Mol Biol Cell. 2016. PMID: 27466320 Free PMC article.
-
The Membrane-Bound Notch Regulator Mnr Supports Notch Cleavage and Signaling Activity in Drosophila melanogaster.Biomolecules. 2021 Nov 10;11(11):1672. doi: 10.3390/biom11111672. Biomolecules. 2021. PMID: 34827670 Free PMC article.
-
Development of the embryonic and larval peripheral nervous system of Drosophila.Wiley Interdiscip Rev Dev Biol. 2014 May-Jun;3(3):193-210. doi: 10.1002/wdev.135. Epub 2014 Apr 16. Wiley Interdiscip Rev Dev Biol. 2014. PMID: 24896657 Free PMC article. Review.
-
Oriented divisions, fate decisions.Curr Opin Cell Biol. 2013 Dec;25(6):749-58. doi: 10.1016/j.ceb.2013.08.003. Epub 2013 Sep 7. Curr Opin Cell Biol. 2013. PMID: 24021274 Free PMC article. Review.
-
A single-cell atlas of the sexually dimorphic Drosophila foreleg and its sensory organs during development.PLoS Biol. 2023 Jun 28;21(6):e3002148. doi: 10.1371/journal.pbio.3002148. eCollection 2023 Jun. PLoS Biol. 2023. PMID: 37379332 Free PMC article.
References
-
- Banks S.M.L., Cho B., Eun S.H., Lee J.-H., Windler S.L., Xie X., Bilder D., Fischer J.A. 2011. The functions of auxilin and Rab11 in Drosophila suggest that the fundamental role of ligand endocytosis in notch signaling cells is not recycling. PLoS ONE. 6:e18259 10.1371/journal.pone.0018259 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
